
Additional Links
| Functional Module: Myosin-X in the Transport of Cargo and Filopodia Initiation
Filopodia are motile structures that
contribute to the cell’s ability to detect mechanical or chemical
signals and measure and respond to its physical environment. This
mechanosensing is mediated primarily by adhesions and receptor molecules
located at the tips, or along the shafts, of filopodia. |
|
|
|
Current MCMF Content |
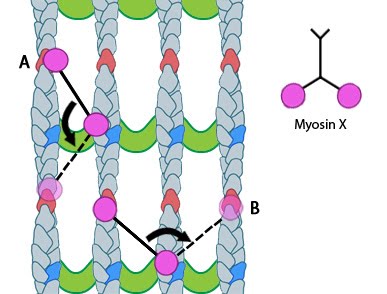
Functional Module: Myosin-X in the Transport of Cargo along Actin Filaments |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites