Read Further…
For a complete introduction to the role of actin in mechanotransduction see: How Actin Filaments Produce, Sense and Convert Cellular Force’
Actin is an essential component in Filopodia, Lamellipodia and Lamellum, Podosomes and Adherens junctions
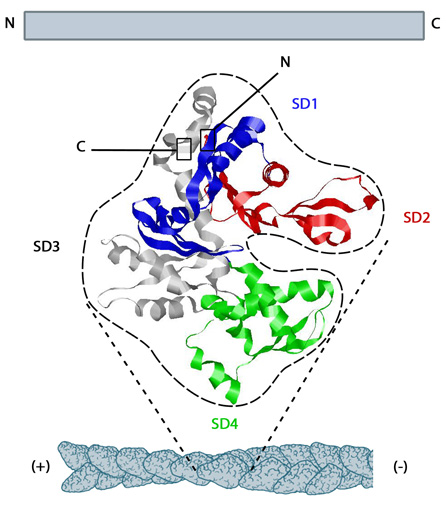
Actin is an abundant (10-100 micromolar on average),~42 kDa structural protein found in all eukaryotic cells (except for nematode sperm). With more than 95% conservation in the primary structure, actin is one of the most highly-conserved proteins [1]. G-actin is the monomeric, globular form of actin that forms the basic subunit for actin filaments. It is believed to fold into an elongated structure that consists
of a large and a small domain which together form a cleft that binds ATP [2, 3].
Polymerization
Actin monomers are assembled to form actin filaments or ‘thin filaments’.
The polymerization of actin filaments, requires ATP-bound G-actin. Assembly of G-actin-ATP is favored at the (+) end of actin filaments. Hydrolysis of ATP to ADP and the steady release of Pi on G-actin cause a conformational change which favors the disassembly of G-actin-GDP at the (-) end of actin filaments. The main actin-binding proteins in vertebrate cells, profilin
and thymosin-β4, control the available pool of monomeric G-actin and
inhibit spontaneous nucleation of actin filaments. Although actin bound
to thymosin-β4 fails to polymerize, profilin can compete with thymosin
for actin binding and can shuttle actin to the barbed-end of a filament [4].
Early models for the actin filament were constructed by fitting the filament x-ray crystal structure to the atomic structure of actin monomers [5] (reviewed in [6]) while more recent models use a number of different approaches [7, 8]. Collectively these results suggest that when single actin strands form, two asymmetric actin monomers align to form a twofold axis of symmetry [9]; their subsequent assembly into a filament that is composed of a pair of strands causes a left-handed helical twist when the adjacent subunits are positioned with respect to each other [10].