
Additional Links Modulators of integrin activation Essential for: Focal adhesion (FA) initiation Podosomes Lamellipodia Related links: Integrins Talin | Functional Module: Activators and Integrin
Integrin activation is an important mechanism through which cells
regulate integrin function by manipulating the ligand affinity of
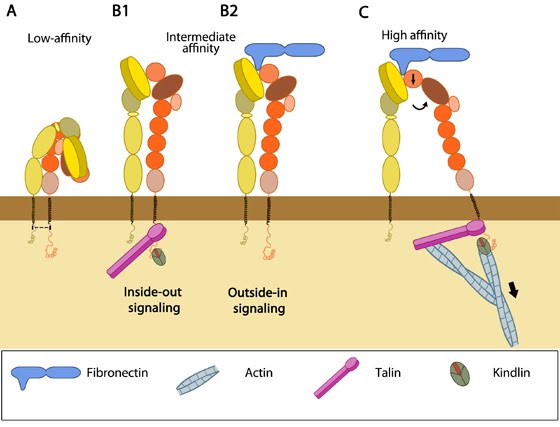
integrins spatially and temporally. Structural and functional studies suggest that integrins can exist in different ligand affinity states- low, intermediate and high (reviewed in [1]). Crystal
structures have revealed that integrin heterodimers,
occur in an inactive, bent V-shape with the head close to the
membrane-proximal regions of the legs [2, 3], maintained by the α/β salt bridge at the inner membrane region and helix packing of the transmembrane (TM) region [4]. This low affinity structure undergoes rapid,
reversible conformational changes to increase ligand affinity, termed “activation” (reviewed in [5, 6, 7]). |
|
|
|
Current MCMF Content |
Functional Module: Activators and IntegrinČ |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites