
Unit 5: Cell-Matrix Adhesions Functional Modules
Additional Resources Test Your Knowledge Cell-Matrix Adhesions Quiz | Cell-Matrix Adhesions5.4 Initiation of Focal Adhesion Assembly
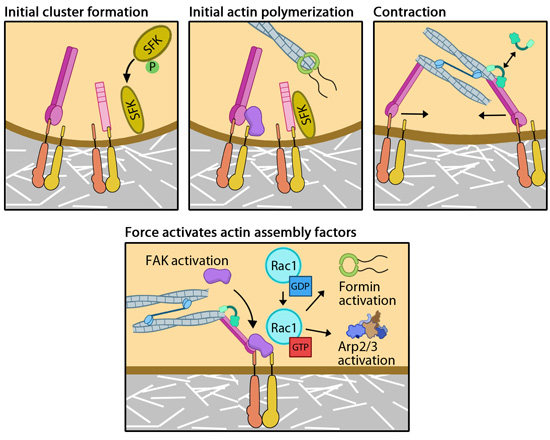
Focal adhesion formation is initiated upon the binding of adhesion receptors to extracellular matrix (ECM) ligands (e.g. fibronectin, vitronectin, collagen) along the cell periphery usually at the protruding edge of a cell. Both intracellular and extracellular factors can influence the level of matrix binding, in terms of affinity (the strength of interactions, reviewed in [1, 2]) and avidity (the number of interactions, such as lateral interactions between independently activated proteins within a focal adhesion). Nascent focal adhesions first appear exclusively in the lamellipodium as submicron-sized puncta that are typically immobile but can at times travel short distances along the direction of the actin retrograde flow [3, 4].
|
|
|
|
Current MCMF Content |
MBInfo > Topics > Cellular Structures in Mechanosensing and Cell Motility > Cell-Matrix Adhesions > Focal adhesions (FAs) >
Focal Adhesion InitiationČ |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites