
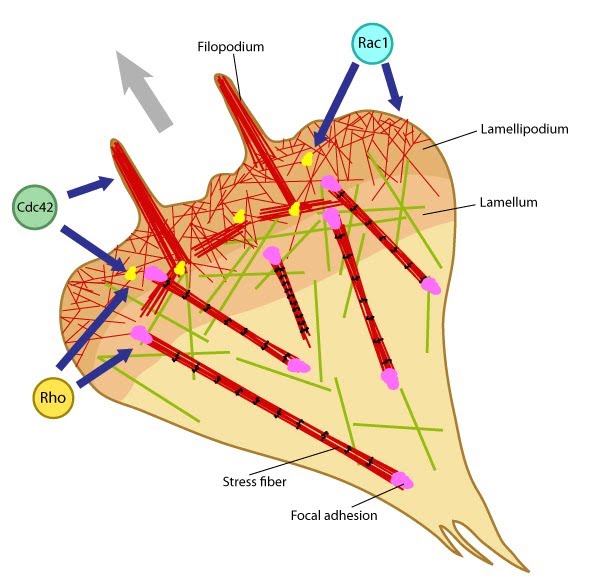
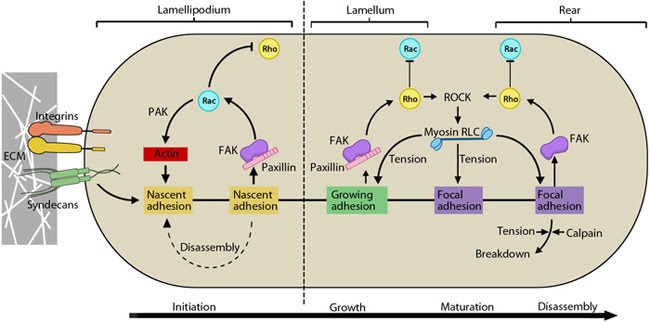
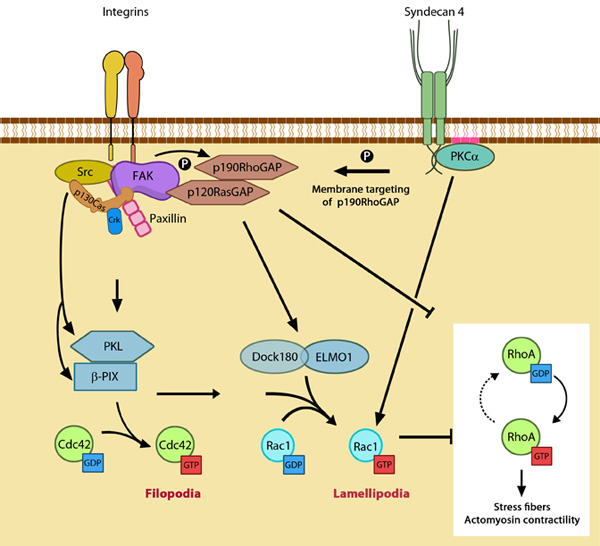
Additional Links Rac activation and Rho suppression are essential for: Focal adhesion initiation Focal adhesions growth Lamellipodial translocation Cdc42 activation and Rho suppression are essential for: Filopodia adherence Rho activation is essential for: Focal adhesion maturation Retraction of the trailing edge and Focal adhesion disassembly Podosome disassembly Cell polarity Influenced by: Guidance signals | Functional module: Integrin β1-syndecan-4 synergy in signaling protrusion and adhesion dynamics
Neither syndecan nor integrin is capable of independently supporting cell adhesion or spreading. Despite the cooperativity of integrin-syndecan pairs in various contexts (reviewed in [1]), recent studies have established synergistic signaling by integrin β1 and syndecan-4; they play cooperative yet distinct roles in cell spreading and maturation of adhesions as well as directional migration respectively [2, 3]. The receptors co-localize in early adhesion sites at the leading edge with ligand binding by both receptors (e.g fibronectin binds via cell binding domain [RGD] to integrin and via HepII domain to syndecan) being necessary for downstream signaling [4, 5]. This is crucial as the cell polarity and migration is determined by differentially regulating signals at the leading and trailing edges. |
|
|
|
Current MCMF Content |
Functional Module: Integrin β1-syndecan-4 synergistic signalingČ |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites