Read Further…
Cyclic Cdc42/Rac and Rho activation during cell motility is mediated by: Integrin β1 and syndecan-4
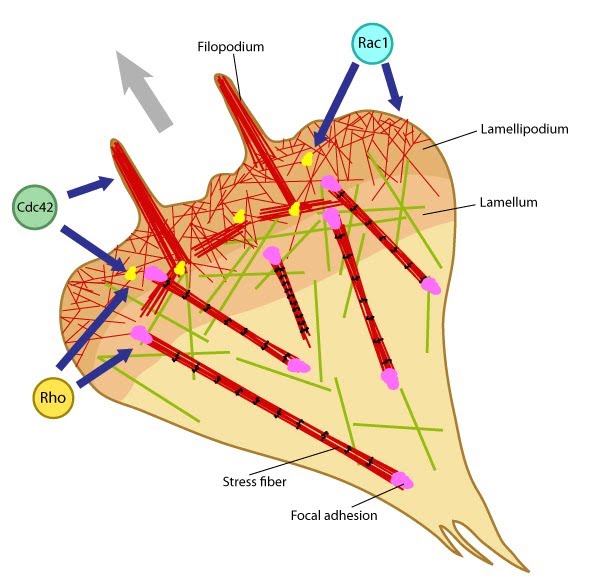
Rac activation and Rho suppression are essential for: Arp2/3 Complex in Actin Branch Nucleation; Focal adhesion initiation and elongation; Lamellipodial translocation
Cdc42 activation and Rho suppression are essential for: Filopodia adherence
Rho activation is essential for: Focal adhesion maturation; Retraction of the trailing edge and Focal adhesion disassembly; Podosome disassembly; Cell polarity
Influenced by: Guidance signals – Attractive cues and Repulsive cues; Activators and Integrin
Rac activation and Rho suppression are essential for: Arp2/3 Complex in Actin Branch Nucleation; Focal adhesion initiation and elongation; Lamellipodial translocation
Cdc42 activation and Rho suppression are essential for: Filopodia adherence
Rho activation is essential for: Focal adhesion maturation; Retraction of the trailing edge and Focal adhesion disassembly; Podosome disassembly; Cell polarity
Influenced by: Guidance signals – Attractive cues and Repulsive cues; Activators and Integrin
The Rho (Ras-homologous) family are Ras-related small GTPases that use the energy from GTP hydrolysis to modulate and control numerous aspects of actin filament dynamics and cytoskeleton structure. Rho GTPases link many cytoplasmic signaling effectors to the actin cytoskeleton [1] (reviewed in [2, 3]) and Rho GTPase activity influences diverse processes such as cell growth, migration, cell shape and cell fate (reviewed in [3, 4, 5]). The specific mechanical and chemical cues that modulate the activity of the more prominent family members, Rho, Rac, and Cdc42, to each of the above mentioned processes is likely to vary between different cell contexts, cell or tissue types, receptors, or stimuli. Although Rho GTPases integrate many signaling events by their interaction with multiple components, these GTPases may act more as permissive factors for cytoskeleton reorganization rather than as direct mediators (reviewed in [6]).
The Rho family members contribute to higher-order actin-based structures such as stress fibers, lamellipodium, dendritic spines, and filopodia [1]. The spatial localization of these GTPases is controlled in different regions of the cell because in certain cases, the activity of one Rho family member antagonizes the activity of another family member (e.g. Rac1 antagonizes RhoA signaling [7]). Conversely, activation of one family member (e.g. Cdc42) can also lead to the stepwise activation of other members (e.g Rac, Rho) [1]. Thus, this significant feedback and crosstalk between the Rho family members complicates the task of constructing one paradigm or linear pathway for Rho-mediated mechanotransduction (reviewed in [8, 9]).
Rac
Rac GTPase activity is primarily associated with actin dynamics and actin-based structures in the lamellipodium (including membrane ruffles). Rac associates with enzymes (e.g. phosphatidylinositol 4-phosphate 5-kinase [10]) that produce secondary signaling molecules such as PIP, which in turn binds directly to several actin-regulating proteins to modulate their activities (reviewed in [3]). Rac GTPase activity also stimulates Arp2/3 complex-mediated assembly of actin filaments through the WAVE family of NPFs [11] (reviewed in [12]); however, this activity can be altered to drive filopodia and growth cone collapse [13]. Rac1 is activated by integrin-ECM binding [14] and Rac1 activity is required for the initial formation of integrin-dependent adhesions in growth cone lamellipodial and filopodial protrusions [15].Rho
Rho GTPase activity is primarily associated with the formation of stress fibers and contractile bundles; its activity influences myosin light chain kinase (MLCK), which in turn, causes increased myosin activity and contraction. Rho members RhoA and RhoB are enriched at nascent adhesion sites [16] and the RhoA effector, Rho kinase (aka ROCK), is necessary for maturation of focal adhesion and myosin-II based contraction in fibroblasts [17]. ROCK activity also promotes rapid neurite outgrowth through stabilization of lamellipodial and filopodial membrane protrusions and maturation of adhesion sites [15]. Lastly, Rho activity removes the autoinhibition of formin dimers to promote their actin nucleating activity and the formation of unbranched actin filaments [18]; a low level of local RhoA activity may be needed for growth cone migration [19].Cdc42
Cdc42 is an important organizer of filopodial dynamics (reviewed in [20]) that uses divergent and separate signaling events to control filopodia activity and growth cone turning [21]. Cdc42 frequently initiates actin assembly via the Arp2/3 complex and WASp. Although Cdc42 appears to be critical for determining cell polarity, Cdc42 activity is redundant in certain contexts [22]. For instance, genetic deletion of Cdc42 abolishes filopodia in primary mouse embryonic fibroblasts, but not in embryonic stem cell-derived fibroblastoid cells [23]. Several other Rho GTPases (e.g. TC10, Rac1, Chp, Wrch-1, RhoD and Rif) also induce filopodium formation through similar effector systems when activated (reviewed in [3]).Regulation of Rho GTPase activity:
Rho GTPases cycle between inactive GDP-bound and active GTP-bound conformations; the activation state is controlled by GAPs (reviewed in [24]), GEFs [25, 26], and GDIs (reviewed in [27]). GTP-binding also influences the cellular localization of the Rho GTPases, with the GDP-bound state existing solely in the cytoplasm due to their association with GDIs. Cellular signals (e.g. growth factors) influence the activity of GEFs at the plasma membrane, which in turn, catalyze the exchange of GDP for GTP on Rho GTPases, thus promoting their activation. Although Rho GTPases form homophillic dimers in both the GTP- and GDP-bound state, homodimerization of only the GTP-bound form causes a significant increase in GTPase activity [28]. The Rho GTPases stimulate actin filament assembly by helping the WASp family overcome their inhibition in cooperation with membrane phospholipids such as phosphatidylinositol 4,5-bis-phosphate (PIP2) [29, 30, 31].