
Cellular Structures
Unit 1: Lamellipodia and Lamella
Unit 2: Filopodia Unit 3: Podosomes Unit 4: Invadopodia Unit 5: Cell-Matrix Adhesions Mechanosensors
Cell-cell adhesion molecules
Cell-matrix adhesion receptors
Contribute | Cellular Structures in Mechanosensing and Cell MotilityCells sense their environment through the detection of forces and the measurement of mechanical stimuli, in a process termed mechanosensing. This process is intertwined with cell motility, which in itself facilitates the ability of a cell to sense its surroundings, but also acts as a response to the detection of a stimulus. In this topic we address the complexities of mechanosensing and cell motility by describing the key structures that facilitate these processes (listed below). We explore how these structures carry out their functions and the common machinery that makes this possible.Contents:Unit 1: Lamellipodia and LamellaUnit 2: Filopodia Unit 3: Podosomes Unit 4: Invadopodia Unit 5: Cell-Matrix Adhesions Unit 6: Cell-Cell Adhesions —————- What are the Key Cellular Structures in Mechanosensing and Cell Motility?Cell polarization, changes in cell shape and cell movement can all be thought of as the end result of local mechanotransduction events, orchestrated into a mechanoresponse at the whole cell level. It is however important to note that cells also use spreading and motility as a means of mechanosensing. Such movements mobilize mechanosensors along the cell periphery to measure changes in the external environment.These movements are reliant on the organization of the intracellular architecture (i.e. cytoskeleton), as well as the protrusive and retractile activities of the membrane, along a particular path. This requires the generation of internal forces, but is also modulated by the detection of external forces. Forces that promote cell motility are primarily propagated through the production of actin-based structures (see Figure below) [1] (reviewed in [2, 3]). These higher order structures include lamellipodia, filopodia, invadopodia and podosomes, which are themselves are inherently motile. The motile behavior of lamellipodia and filopodia is dependent on the turnover of cell-matrix and cell-cell adhesions. 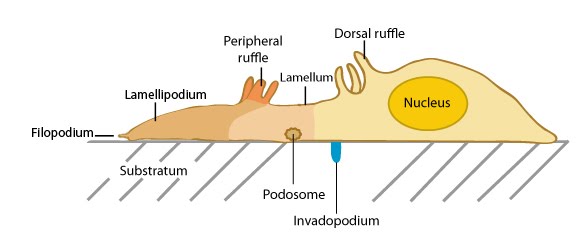 Interactive Figure: Actin-based motility structures. Click on the structure name for more information on its location and function. Actin filaments are a basic structural component found in a number of cellular structures used for cell motility in metazoan cells (insets: filopodium, lamellipodium and lamellum). Certain structures appear sheet-like (e.g. lamellipodia/lamellae, ruffles), whilst others are finger-like (e.g. filopodia, podosomes). In the top panel, the cell is migrating upwards and is attached to a second cell on the right. The bottom panel shows the side view of the migrating cell. Although migrating cells come in many different forms, the leading edge is dominated by the lamellipodium. This structure is highly conserved across cell types and is the key structural element that drives cell migration and spreading. Furthermore, the protein constituents that contribute to lamellipodium formation and maintenance are conserved in many other actin-based extensions (reviewed in [4]). It can therefore be said that the motile behavior of cellular structures, including lamellipodia, filopodia, invadopodia and podosomes, occur through the action of common protein complexes, which are described in this resource as Functional Modules. Additional cytoskeleton components (e.g. microtubules) are important for actin dynamics and for establishing the deposition pattern of the actin network, which both determine the polarity of cell migration (reviewed in [5, 6, 7]). The specific contribution of these components will be discussed throughout. What are the Functional Modules that Mediate Mechanosensing and Cell Motility?In the case of cell motility, each functional module acts to regulate the dynamic state of the cytoskeleton. By viewing cell motility as the sum of all the functional modules that are active at a given time, one can reason that different combinations of active modules will allow the cell to achieve a large number of functions from a limited set of proteins. This results in a variety of different migratory and mechanosensory behaviors.The Functional Modules that mediate motility, either of specific structures or of the entire cell, are:
How are Mechanical Forces Detected?The detection of mechanical force is facilitated through proteins or protein complexes that form mechanosensors. In response to a change in force, mechanosensors commonly exhibit structural rearrangements or geometry-dependent clustering that promote biochemical and mechanotransduction events (reviewed in [8]). These changes in response to force fall into two broad categories:
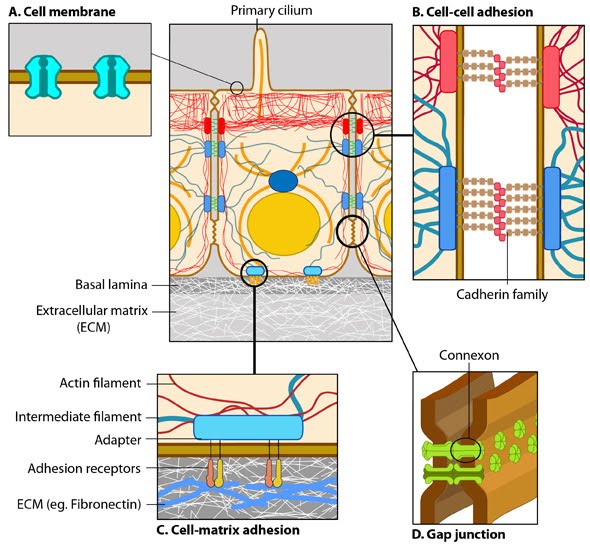 |
|
|
|
Current MCMF Content |
MBInfo >
Cellular Structures in Mechanosensing and Cell Motility |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites