
Motor Proteins in Mechanobiology4.3 An Example of Function: A Role in ContractionAs described in the previous chapter, numerous motor proteins exist and each possess unique characteristics that allow them to facilitate different processes and functions. Even within the myosin superfamily variation exists in structure and function of each member.One example of a process in which the motor proteins are involved is in the formation and function of contractile bundles. Contractile Bundles in Skeletal MuscleIn skeletal muscle cells, myosin II forms only thick filaments and they are arranged within a scaffold of actin thin filaments (along with numerous other proteins) into higher order fibrous structures known as sarcomeres. Each sarcomere contains numerous repeating units of interlinked thick and thin filaments, and the opposite orientation of the myosin heads causes adjacent actin filaments to slide past each other during muscle contraction. Each sarcomere is ~2 µm long in resting muscle, but this length is shortened by as much as 70% after muscle contraction. Muscle contraction is regulated by calcium levels [1] and by the troponin regulatory system (see figure “Tropomyosin stabilizes thin filaments“). Although actin subunits continue to turn-over at both ends of the thin filament, this exchange is relatively slow, making the actin filaments in sarcomeres relatively more stable when compared to the actin filaments found in other cell types.Video: Sliding filament theory of muscle contraction. [Video was uploaded to YouTube by avice01 and created by Sara Egner using electron micrography from P M Motta, P M Andrews, K R Porter and J Vial.] 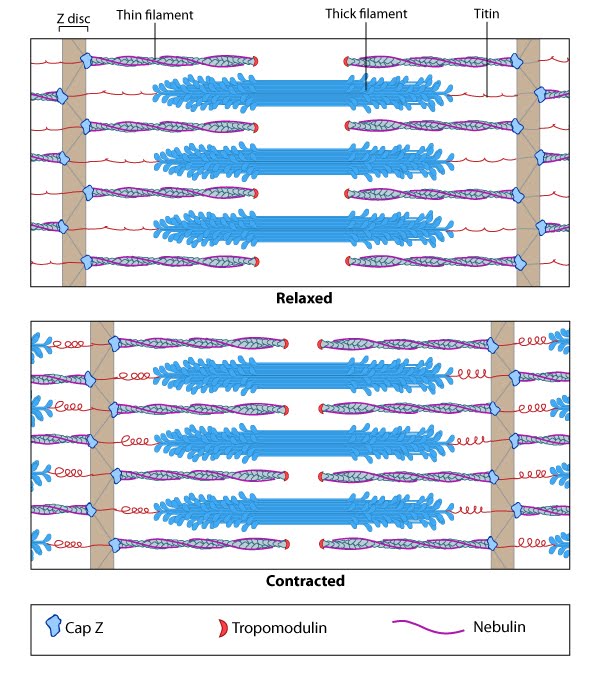 Contractile Bundles in Nonmuscle CellsIn nonmuscle cells, myosin II associates with actin filaments to form contractile structures known as stress fibers along the lower surfaces where the cell is anchored to its substrate. In epithelial cells, contractile bundles are also prominent in the adhesion belt (aka adherens belt; see also “adherens junction“), which helps to maintain the stability and integrity of epithelial cell sheets. The contractile bundles in nonmuscle cells are similar to skeletal muscle fibers, but they are smaller (~0.4 µm in fibroblasts), less organized, and they contain different accessory proteins [2]. Historically speaking, the mechanism of actomyosin contraction for nonmuscle actin was examined using amoebae proteins Dictyostelium, Acanthamoeba) because the actin is very similar to muscle actin [3]; these initial studies showed the rate of ATP hydrolysis by myosin (and hence myosin movement) varies directly with the actin concentration [4]. Further studies using isolated stress fibers from fibroblasts confirmed that stress fibers are contractile and shorten by as much as 25% [5]. Myosin II bundle formation and contractile activity in nonmuscle cells is regulated by phosphorylation [6].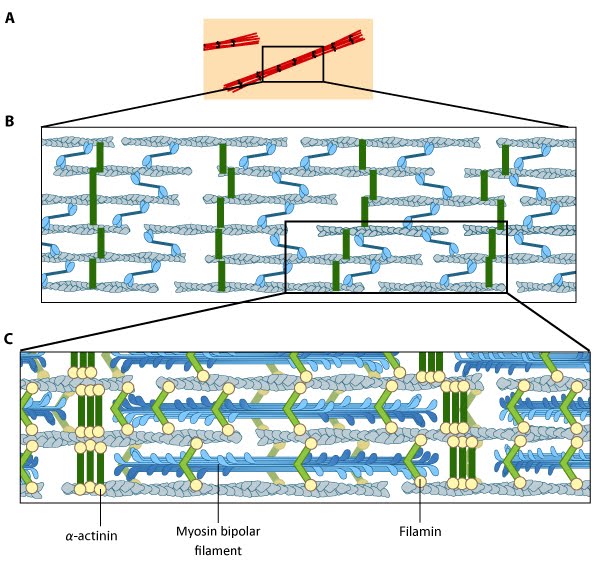 |
|
|
|
Current MCMF Content |
MBInfo > Topics > Mechanobiology: When Cells Sense and Respond to their Environment > What are Motor Proteins? >
Myosin- role in contraction |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites