
Glossary Term: Alpha (α)-catenin
Protein structure:
alpha (α)-catenin is a well conserved member of the actin filament crosslinking functional module and an integral component of cell-cell adhesion complexes (reviewed in [1, 2, 3, 4]). α-catenin has three isoforms in mammals known as αE-,αN- and αT-catenin (E, epithelial; N, neural; T, heart and testis) based on the tissue from which they were originally identified (reviewed in [3, 4]). α-catenin has potential interaction domains for numerous proteins (including itself [5]) (reviewed in [3, 4]) and it can be phosphorylated [6]. α-catenin has three regions in common with vinculin (known as vinculin homology domains [VH1-VH3]) [7] which places these proteins within the same family (reviewed in [3, 4]). α-catenin is a mechanosensor that naturally exists in both monomeric and dimeric forms with each form having their own binding properties and structural conformation [8]. α-catenin can cross link and organize actin filaments directly via its actin binding domain (ABD) located at the C-terminus [8] or through its binding partners (e.g. vinculin [9] and α-actinin [10]). α-catenin also binds to and/or influences components that control the production of actin filaments (e.g. formins, Arp2/3 complex, Ena/VASP) (reviewed in [3, 4]).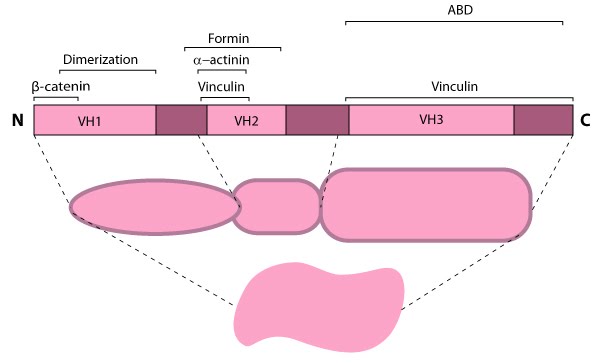
Localization and function
α-catenin has broad activity that contributes to processes such as differentiation (i.e. commitment to a particular cell type), embryonic and tissue development, and cell migration (reviewed in [3, 4]). α-catenin is concentrated at cell-cell adhesion sites, e.g., tight junctions [9], and adherens junctions (reviewed in [2]), through its association with a related family member, beta (β)-catenin; this binding interaction is controlled by phosphorylation of either α- or β-catenin [5, 14] (reviewed in [15]) and phosphorylated β-catenin is expected to compete with homodimerziation of α-catenin [5, 11]. Dimerization of α-catenin creates a complex with functional domains at both ends that preferentially binds actin filaments, in contrast to the monomer which prefers E-cadherin-β-catenin complexes [8].Because β-catenin binds tightly to classical cadherins before they are transported to the cell surface [16, 17]), it was originally suggested that α-catenin formed an indirect link between adhesion receptors and the actin cytoskeleton via its association with β-catenin or other actin-binding proteins (e.g. vinculin and α-actinin) [17]. However, later work showed that α-catenin cannot bind the E-cadherin-β-catenin complex and actin simultaneously, nor does it bind actin indirectly through its binding partners, vinculin or α-actinin [18]. Thus, rather than serving as a bridge to the cytoskeleton, α-catenin appears to function primarily as a molecular switch that promotes stable cell-cell adhesions (e.g. adherens junctions).
Current models suggest that α-catenin promotes stronger adhesions in a few ways: 1) α-catenin may foster lateral clustering and activation of cadherins [19]; 2) α-catenin may recruit formins at nascent cell-cell contacts to produce new filaments that push against and bring the membranes together [13] (reviewed in [3, 4]); and 3) α-catenin may regulate membrane protrusive activity [20] and suppress Arp2/3 complex-mediated actin nucleation and polymerization at the leading edge in the lamellipodium [8] (reviewed in [4]).