
Contents
1.1 Role of Mechanobiology in Shaping Cells and Tissues 1.2 Common Themes In Mechanobiology 1.3 Types of Mechanosensing 1.4 Types of Forces Cells Encounter 1.5 The Dynamic Cytoskeleton 1.6 Common Features of Polymeric Cytoskeletal Systems 1.7 How Does the Cytoskeleton Transmit Mechanical Forces? Contribute | Essential Info: What is Mechanobiology?
|
|
|
|
Current MCMF Content |
Essential Info: What is Mechanobiology? |
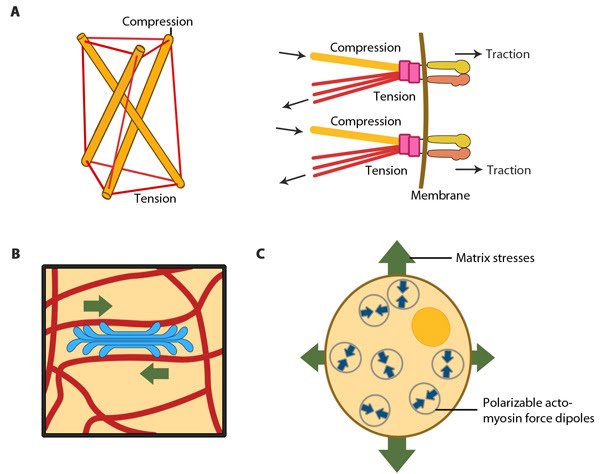
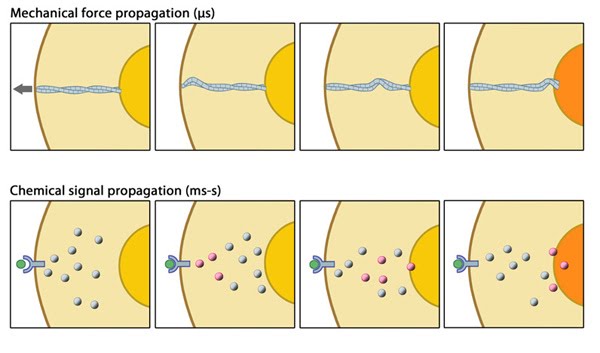
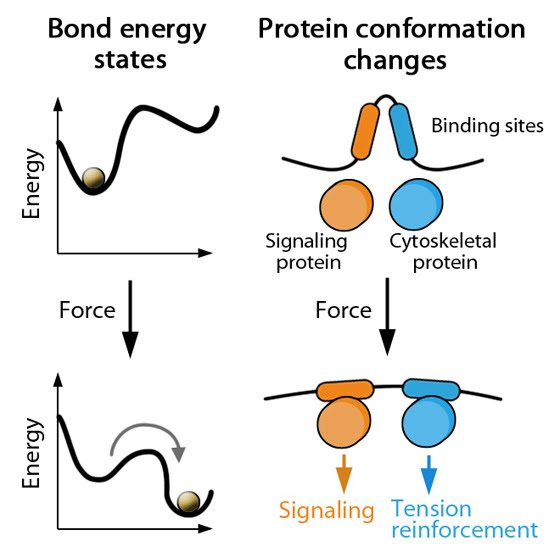
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites