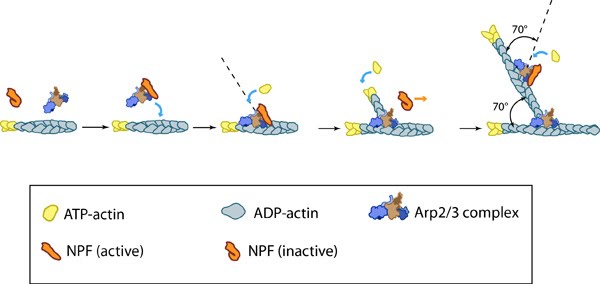
Accessory proteins nucleate actin filaments.: A. NPFs (e.g. WASp; Scar) bring together the Arp2/3 complex and actin monomers to nucleate new actin filaments and to form new branches from the side of pre-existing filaments. Arp2/3 complex remains at the minus end of the filament. B. Formin cooperates with profilin to nucleate new actin filaments. Formin remains at the plus end of the filament.
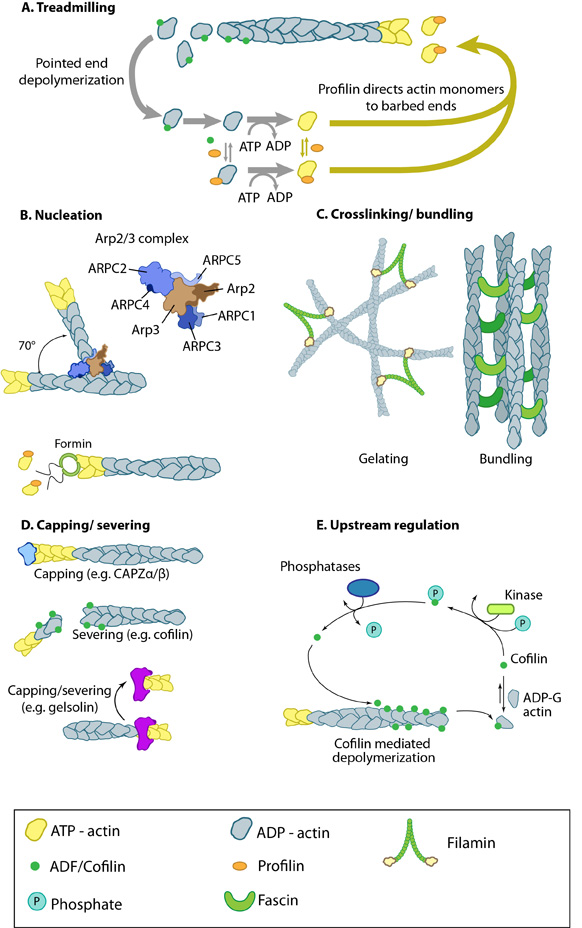
Actin binding proteins influence actin dynamics: A. Treadmilling of actin filaments can be altered by profilin and ADF which generally increase and decrease the size of actin filaments, respectively. B. New filaments are nucleated by the ARP2/3 complex, which binds both G-actin monomers and the side of actin filaments to nucleate new filaments or branches. Formins nucleate new filaments by binding G-actin and through cooperation with profilin. C. Actin cross-linking proteins influence the packing and organization of actin filaments into secondary structures. D. Capping and severing proteins promote disassembly of actin filaments. E. Actin filament assembly can be modulated by events such as controlled nucleotide hydrolysis (e.g. ATP on actin) and reversible modifications (e.g. phosphorylation) on components that control actin assembly.
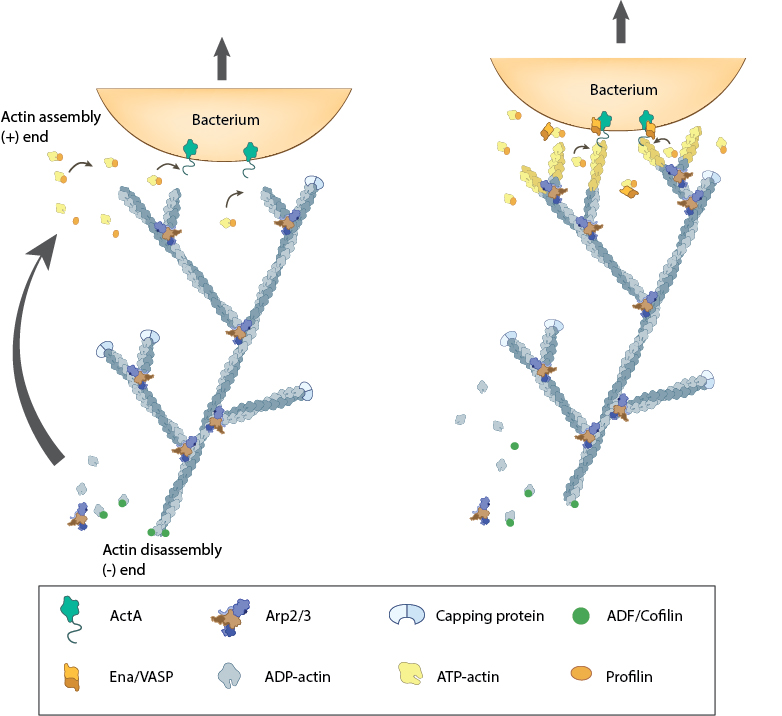
Actin polymerization produces force for movement: Pathogenic intracellular bacteria use mainly host-derived components to facilitate movement through (and between) cells. The bacterial ActA protein on the surface of the bacterium (e.g. Listeria monocytogenes) recruits and activates the host components needed for actin polymerization (e.g. Ena/VASP, ATP-actin). The actin filaments are assembled at the plus end nearest the bacteria membrane and the forces generated by filament assembly are translated into bacterium movement. The resulting actin comet tail is likely comprised of branched arrays of actin filaments as seen from simulation of experimental results [3, 8, 28]. The dynamics of filament assembly and turnover (and therefore bacterium motility) are controlled by capping protein, profilin, ADF/Cofilin and Arp2/3 complex.
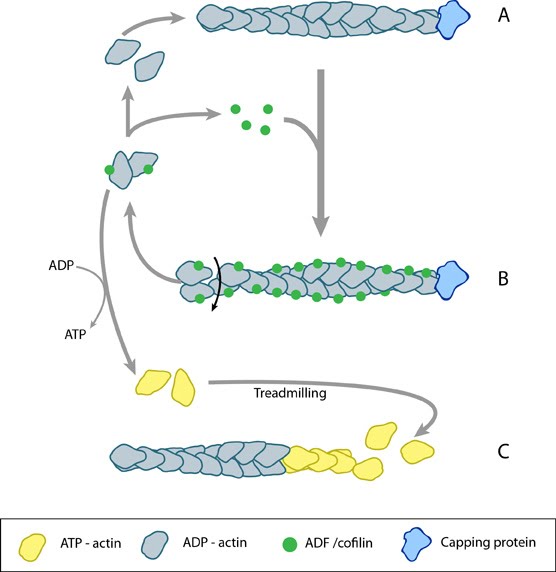
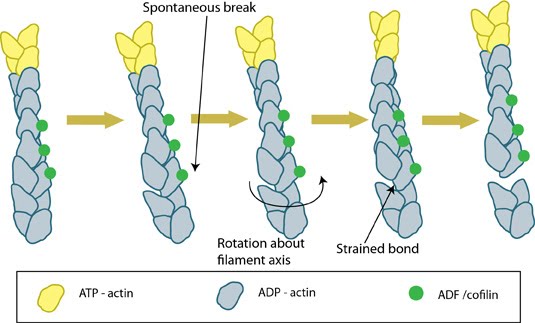
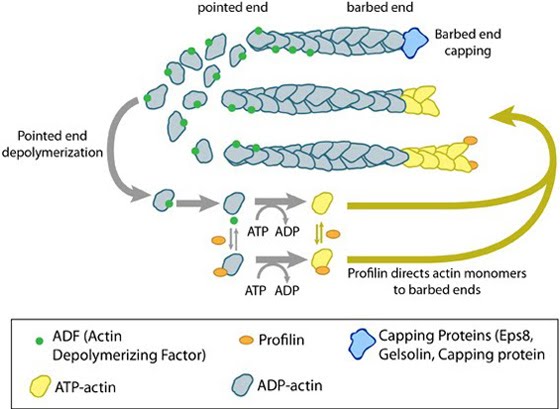
ADF/cofilin influences actin filament turnover: ADF/cofilin influences actin filament turnover. ADF cooperatively binds to F-actin to increase the steady state turn-over (e.g. treadmilling rate) of actin filaments and accumulation of ADF-ADP-G-actin, ADP-G-actin, and ATP-G-actin (via nucleotide exchange). ADF binds faster to actin filaments that have barbed end capping proteins (e.g. gelsolin) (B). The actin subunits depolymerizing from ADF-bound filaments are recycled to filaments lacking ADF, thus maintaining the pool of actin-filaments. At high ADF concentration, polymerization of ADP-actin at the pointed end is favored on capped-filaments (A); alternatively, nucleotide exchange increases the pool of ATP-actin for barbed end assembly of new or existing filaments (C).
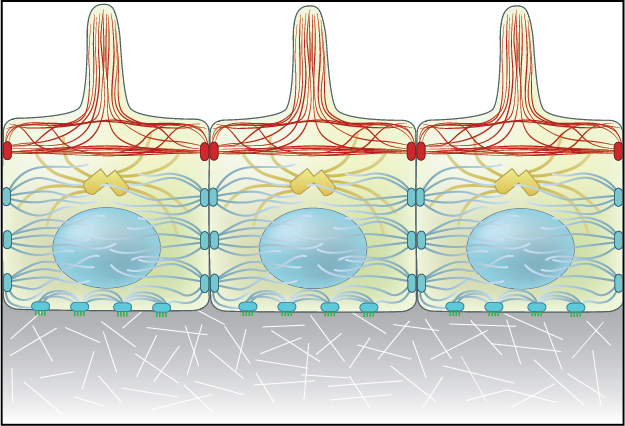
Adherens junctions link actin filaments between cells.: Adherens junctions link actin filaments between cells. This diagram specifically depicts adherens junctions (red rectangles) connecting actin filaments (red lines) across polarized epithelial cells. In these cells, this results in the formation of contractile bundles of actin and myosin filaments near the apical surface – this forms a structure called the adhesion belt (arrows). Other junctions termed desmosomes (large blue rectangles) and hemi-desmosomes (small blue rectangles) link intermediate filaments (blue lines) between cells.
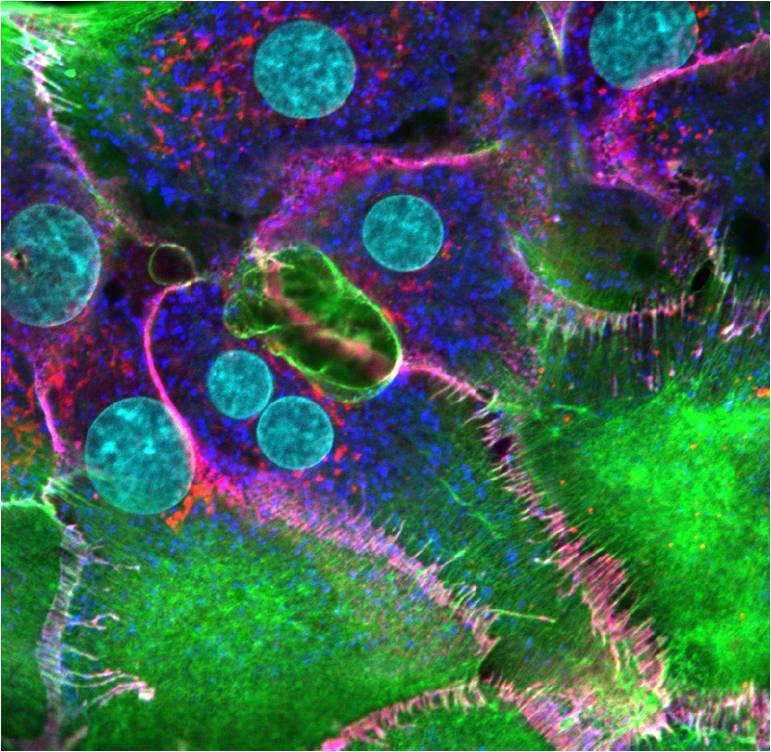
Adherens junctions of hepatocytes: Hepatocyte cells plated on collagen and stained the next day to image newly formed cell-cell contacts. F-actin (green), β-catenin (red), cadherin (dark blue), nuclei (cyan). The overlap of β-catenin and cadherin seen as magenta represents adherens junctions that have formed between apposing cells. These cells were imaged using a Nikon A1Rsi confocal microscope at 100x magnification. [Image captured by Jeffrey Robens, Mechanobiology Institute, Singapore].
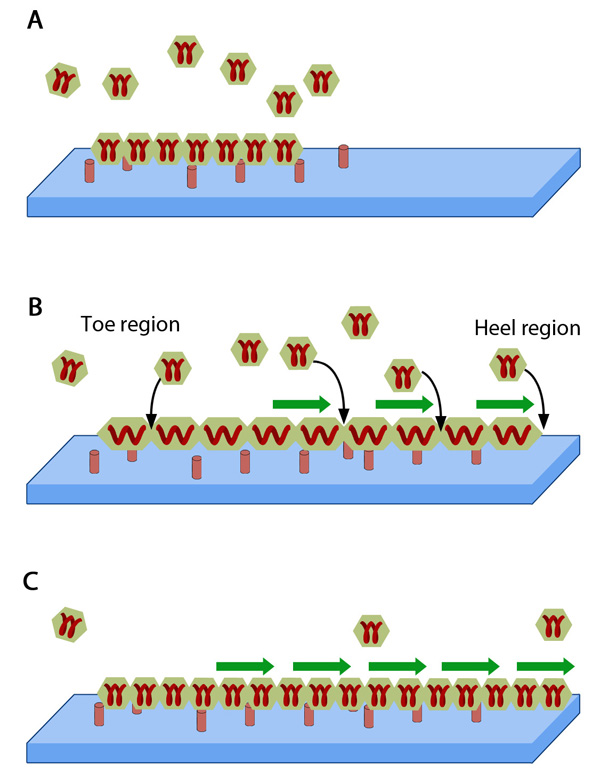
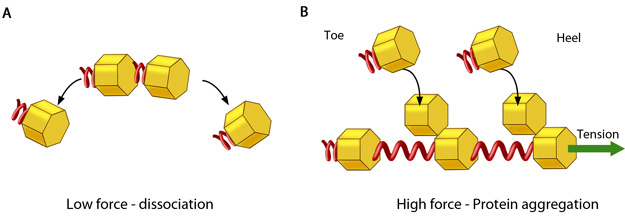
Adhesion growth under force: In the absence of or under low actomyosin contractile forces, adhesion components start dispersing. In such scenarios, the unidirectional shear force (not shown) from the actin retrograde flow may also aid dissociation. B. Under substantial pulling 'tension' from actomyosin contractions, aggregation of adhesion components occurs in the direction of pulling. The protein complexes of adhesions (cell's feet) can be considered as elastic units that expand under force to accommodate new components towards one end (heel). The other end of the adhesion structure that faces the actin retrograde movement is termed 'toe'. Adapted from [18].
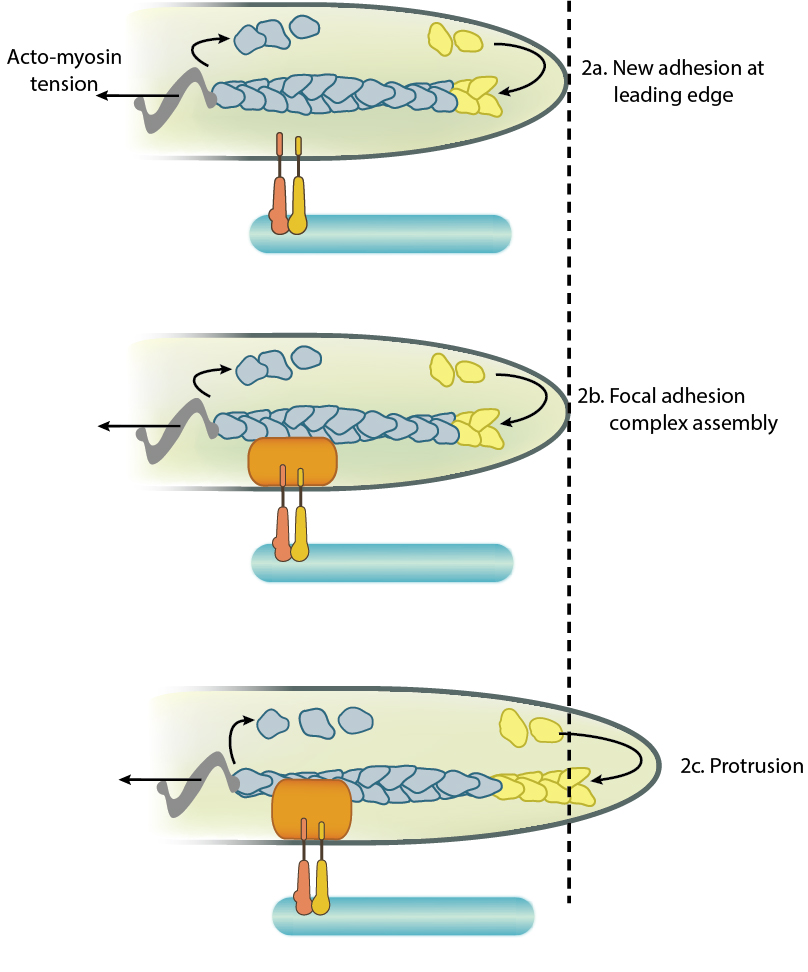
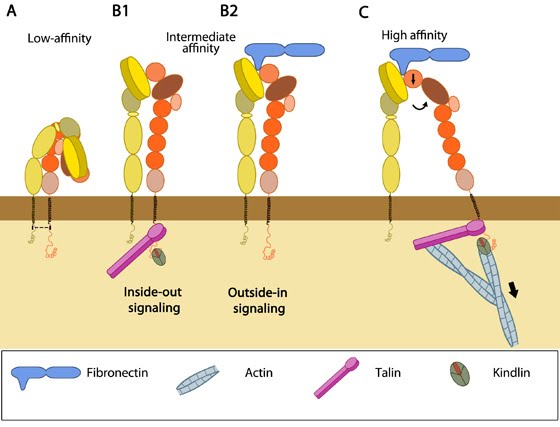
Adhesion influences filopodia protrusion: Adhesion acts like a "molecular grip" and influences the protrusion of filopodia (2a). Adhesion between membrane-anchored integrins and the extracellular substrate (e.g. fibronectin) causes clustering of focal adhesion components (represented by a hand,2b). Connections between the adhesions and the actin cytoskeleton converts the force generated by actin polymerization into protrusion(2c).
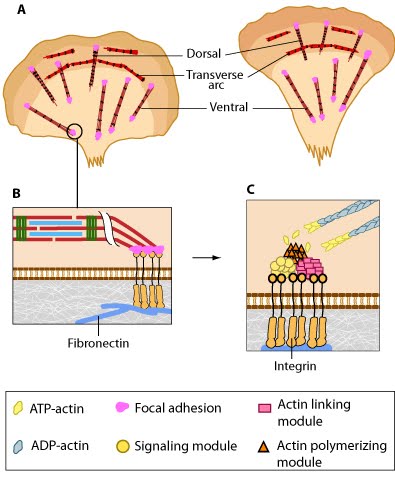
Adhesion of dorsal stress fibers: Soluble clues or binding of cell surface receptors (e.g., integrin) to the substratum components (e.g., fibronectin) promotes clustering of adhesion receptors and subsequent activation of the Rho family of GTPases (e.g., Rac1). Rac1 activity strengthens the nascent adhesion by promoting the assembly of scaffolding proteins and actin-binding proteins to the adhesion site (figure adapted from [16651381])
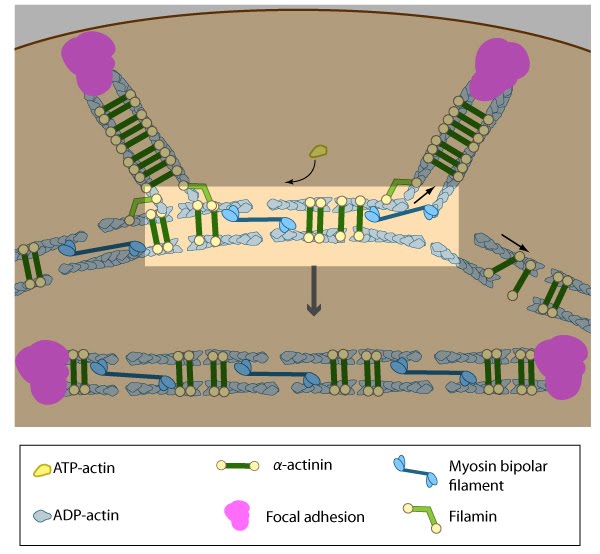
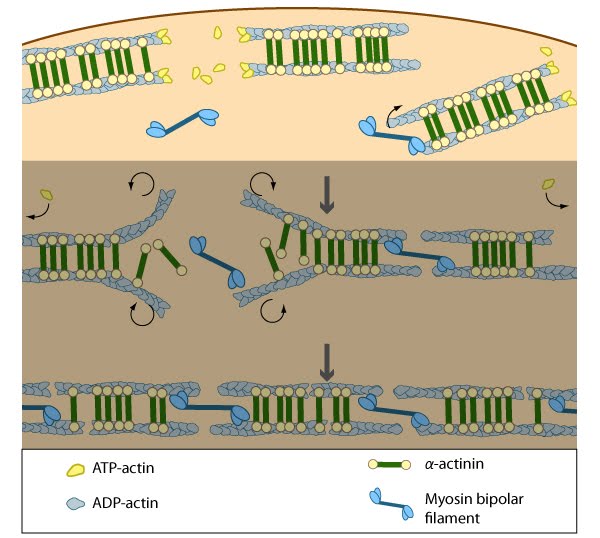
Alignment of ventral stress fibers: The pre-existing dorsal stress fibers interface with the transverse arcs which leaves the arc filaments aligned between the two dorsal stress fibers. Myosin bundles donated by the transverse arc continue to contract, which helps bring the fused structure into complete alignment.
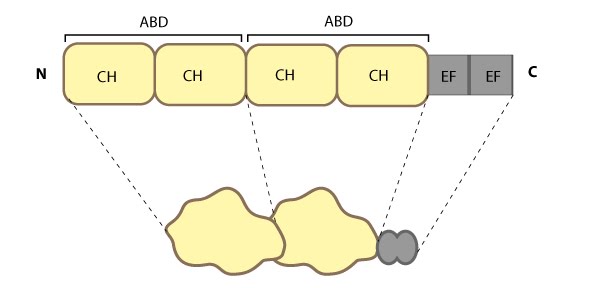
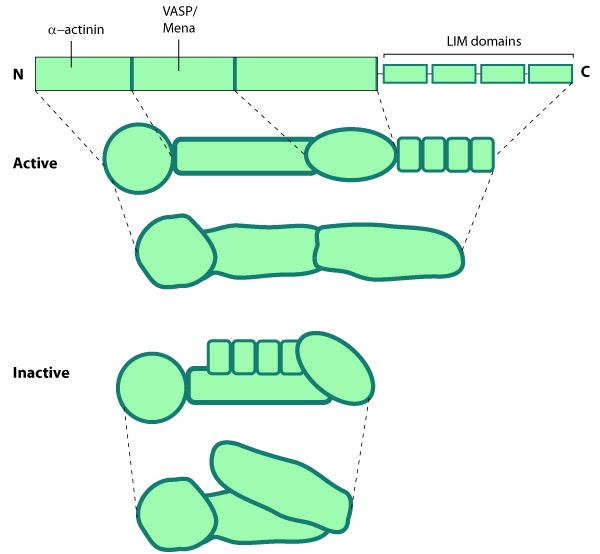
Alpha (α)-actinin: This schematic diagram illustrates the molecular organization of α-actinin (reviewed in [18488141]) and provides examples for how α-actinin is represented in figures throughout this resource. Relevant domains/regions that are believed to be important for actin binding and protein-protein interactions are highlighted (actin binding domain (ABD) [3733725], β-integrin [2116421, 15721583], α-catenin [9152027] and vinculin [15988023, 8037676]).
15.
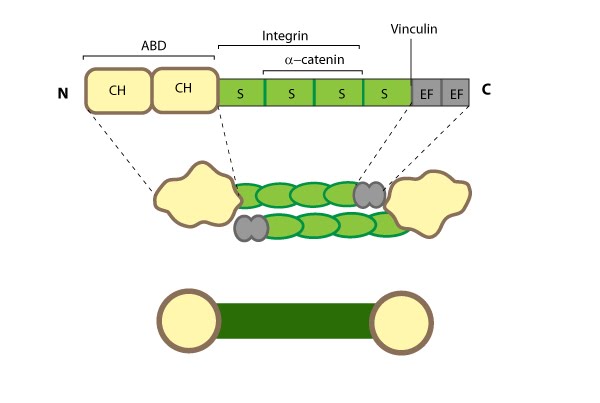
Alpha (α)-catenin: This schematic diagram illustrates the molecular organization of
α-catenin and provides examples for how α-catenin is represented in
figures throughout this resource.
16.
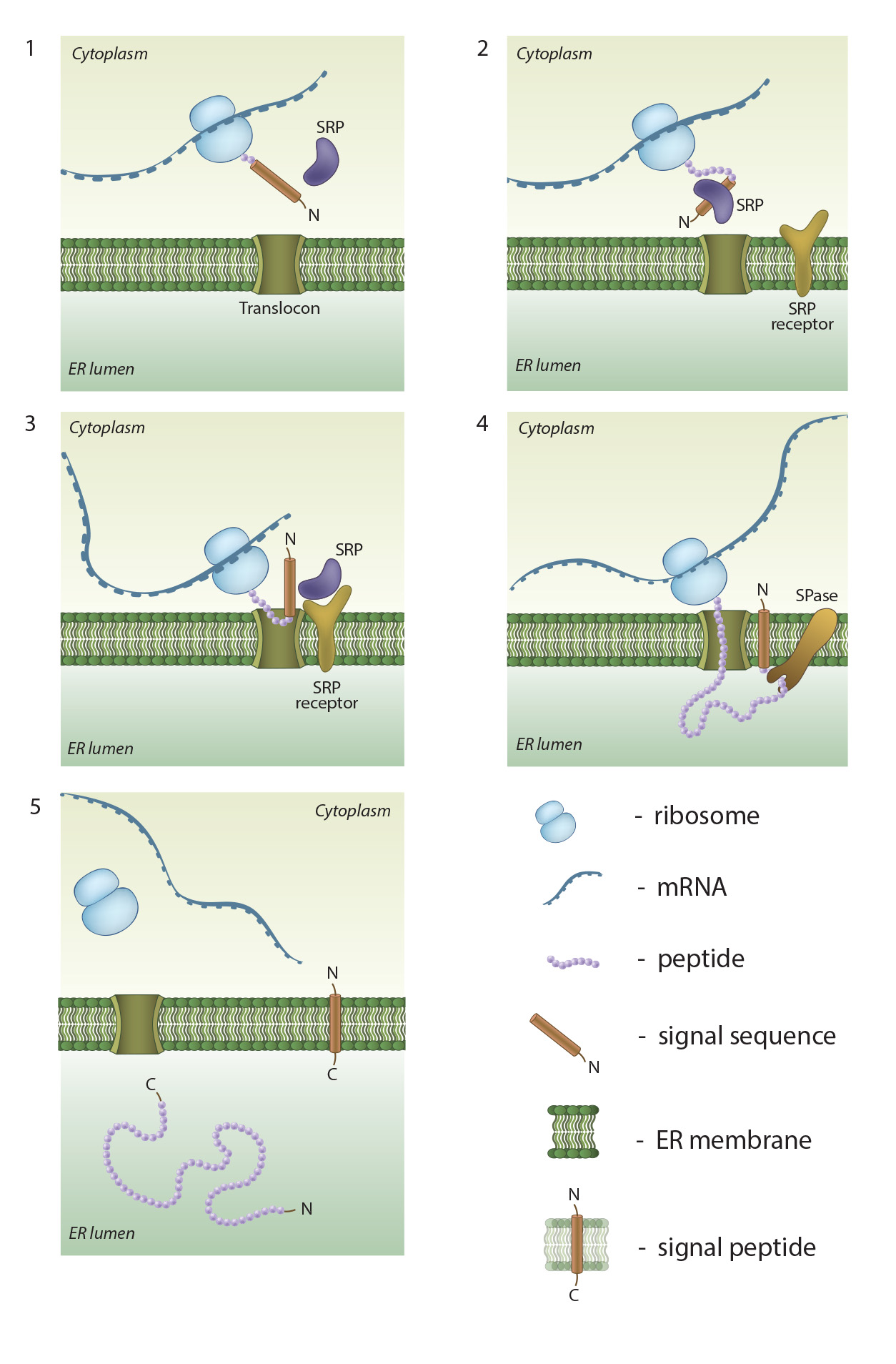
An
overview of co-translational targeting of proteins destined for secretion or
membrane insertion: SRP (signal recognition particle) interacts with the signal
sequence as soon as it emerges from the ribosomal polypeptide exit tunnel (1-2).
In eukaryotes peptide elongation pauses upon SRP / ribosome nascent chain complex
formation; the complex is then targeted to the ER membrane by the interaction
with the SRP receptor (3). GTP binding to SRP and SRP receptor is a
prerequisite for SRP / SRP receptor complex formation. Following the complex
positioning (3), SRP / SRP receptor complex dissociates from the peptide due to
GTP hydrolysis. The growing peptide is gradually transferred via the
protein-conducting channel (the translocon) into the ER lumen (4). Usually, signal sequences are cleaved off co-translationally by
SPases (signal peptidases), and the resulting cleaved signal sequences are
termed signal peptides (4-5).
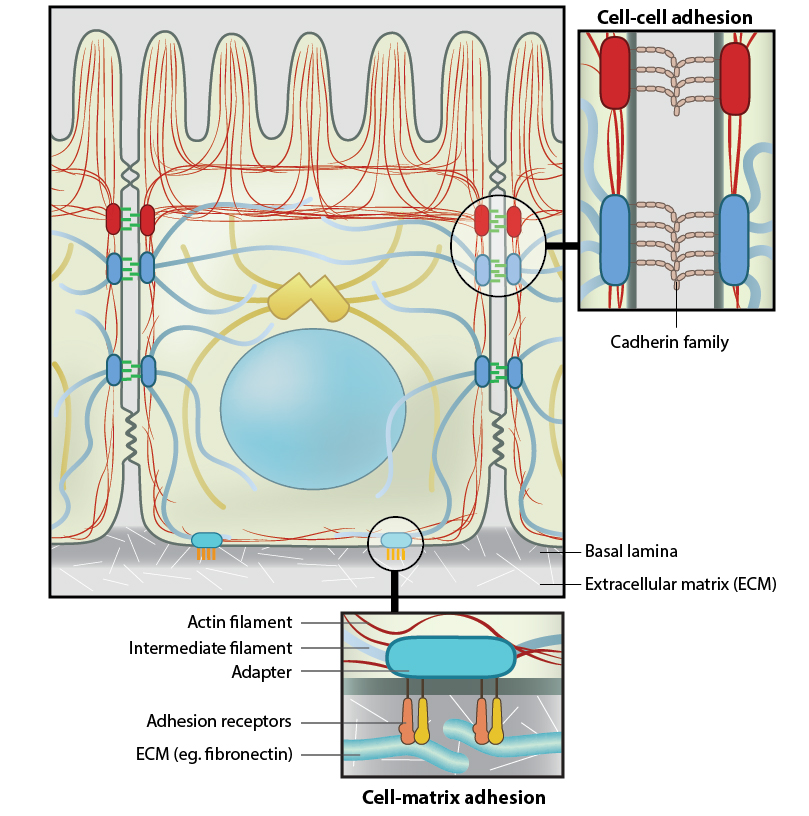
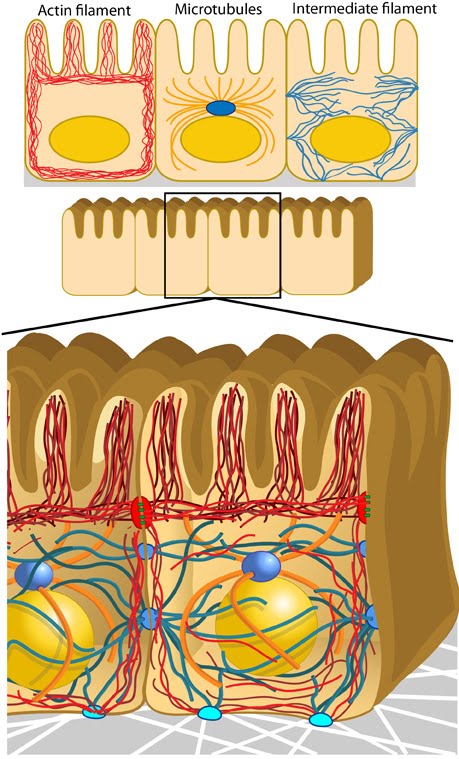
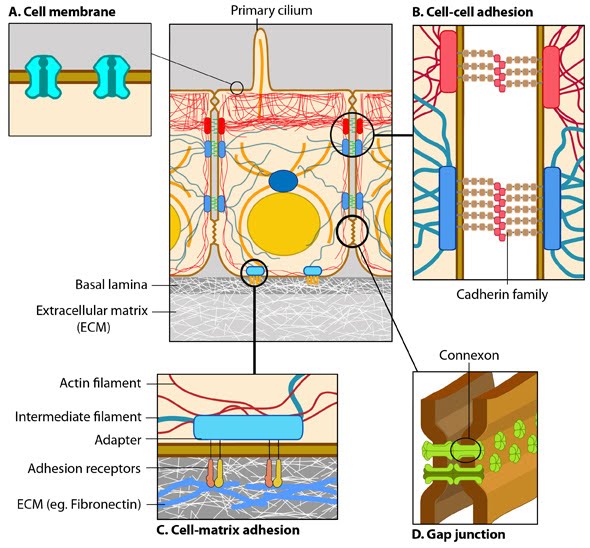
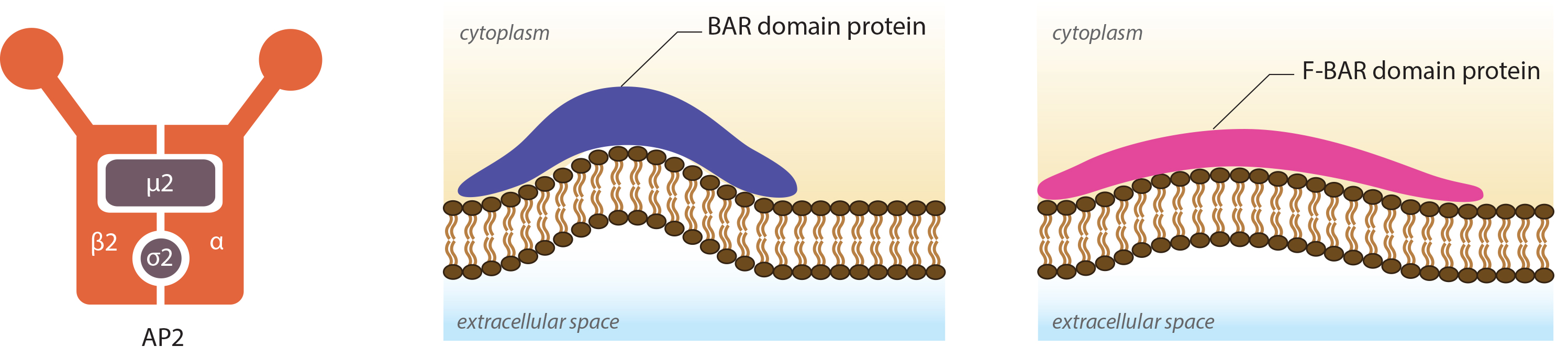
Anchoring junctions: Actin filaments are shown in red, microtubules are yellow, and intermediate filaments are blue. Transmembrane complexes (solid ovals) contain several different components that play a key role in linking the cell exterior to the cell interior. Cell-cell adhesion: Numerous cell-cell adhesion molecules (e.g. cadherin family of proteins) and their associated cytoplasmic anchoring components (e.g. vinculin, α-actinin) form a continuum between cells and their linked cytoskeleton components. Adherens junctions (shown as a solid red oval) primarily link actin filaments between cells, while desmosomes (shown as a solid dark-blue oval) primarily link the intermediate filaments between cells. Cell-matrix adhesion: Interactions at CMACs and hemidesmosomes (shown as a solid light-blue oval) link the actin and intermediate filaments to the underlying matrix, respectively. Cell-matrix linkages are a key force-sensing unit that greatly influences cell polarity and migration
18.
Annealing of transverse arcs: As actin filaments are transported away from the cell edge, they further evolve through end to end annealing and associate with myosin-containing bundles. The filaments undergo continued adjustment and polarity sorting to establish contractile arcs. Actin subunits are exchanged throughout the arcs either as a result of polymerization onto the ends of actin filaments, binding to stress fiber myosin, actin treadmilling or interaction with actin-binding proteins in stress fibers (figure adapted from [16651381])
Arp2/3-mediated actin polymerization: NPFs (e.g. WASp; Scar) bring together the Arp2/3 complex and actin monomers to nucleate actin filaments that form new branches from the side of preexisting filaments. The Arp2/3 complex remains at the minus end of the filament.
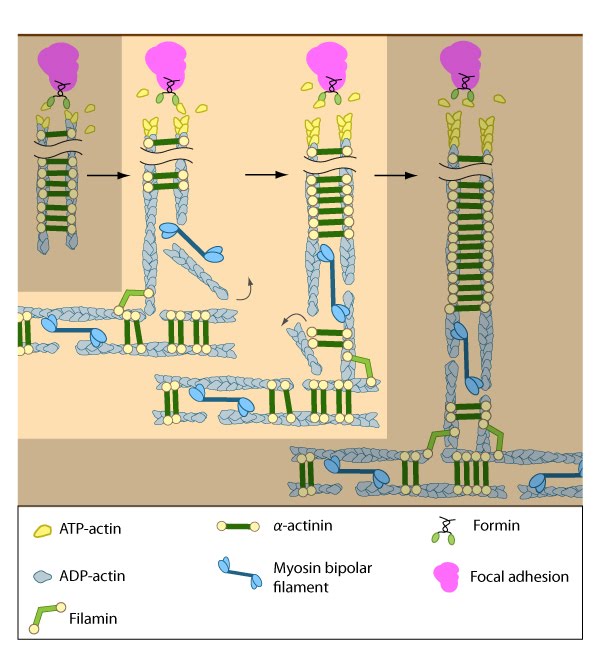
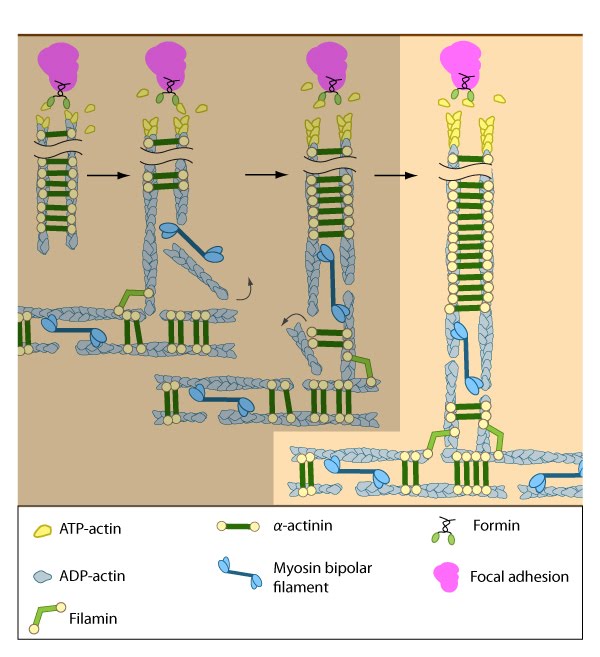
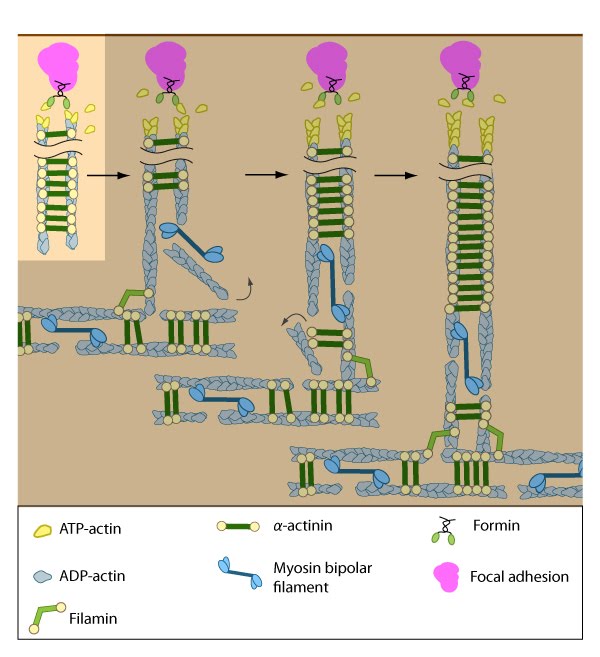
Assembly of dorsal stress fibers: The nascent adhesions continue to grow and exchange components. Nascent unipolar actin filaments are extended and they are stabilized by actin binding proteins (e.g., α-actinin, filamin). The population of filaments are crosslinked into bundles and they steadily exchange actin subunits as they move towards the cell body due to retrograde actin flow and actin treadmilling. The orientation and polarity of the filaments may become mixed as they are transported (figure adapted from [16651381]).
Assembly of transverse arcs: Nascent unipolar filaments are extended and are rapidly bound by actin binding proteins (e.g., α-actinin). The population of filaments steadily exchanges actin subunits and moves towards the cell body due to retrograde actin flow and actin treadmilling; the orientation of the filaments may also change with the direction of movement (figure adapted from [16651381])
Association of ventral stress fibers: Pre-existing dorsal stress fibers interact with transverse arcs to demarcate a region of the arc that will align between the two dorsal stress fibers. Crosslinking of the actin filaments by actin binding proteins assistss this interaction. Actin subunits are exchanged throughout the stress fibers and arcs either as a result of polymerization onto the ends of actin filaments, binding to stress fiber myosin, actin treadmilling or interaction with actin-binding proteins in stress fibers (figure adapted from [16651381]).
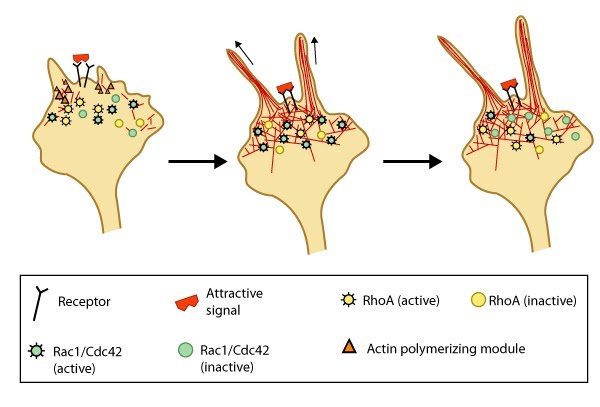
Attractive cues promote filopodia formation: In neuronal growth cones, filopodia protrusion is induced when an attractive cue binds to its receptor on the cell surface. Receptor-binding transiently activates Cdc42/Rac1 GTPase and subsequent recruitment of adhesion molecules and components of the actin polymerizing module (e.g. Arp2/3 complex and formins such as mDia2)which together promote filopodia protrusion. Rac1/Cdc42 activity is then locally reduced in the filopodium, which allows RhoA and its effector, ROCK, to become activated; ROCK is involved with subsequent stabilization and maturation of adhesions, which supports migration in the direction of the attractive signal.
25.
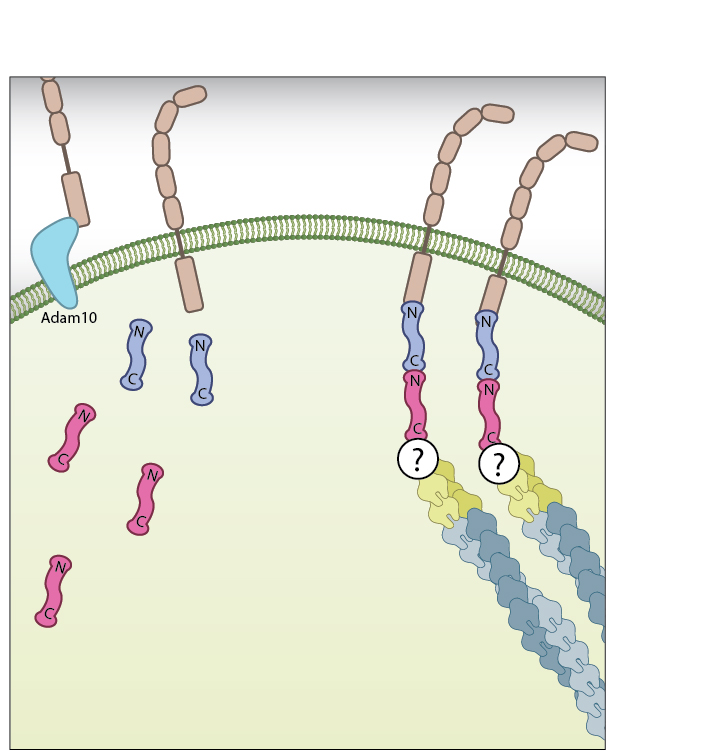
B-catenin at the membrane: β-catenin exists as part of the adhesion complex, bound to E-cadherin and α-catenin, at the cell membrane. Disassembly of the adhesion complex by proteases such as ADAM10 releases β-catenin into the cytoplasm.
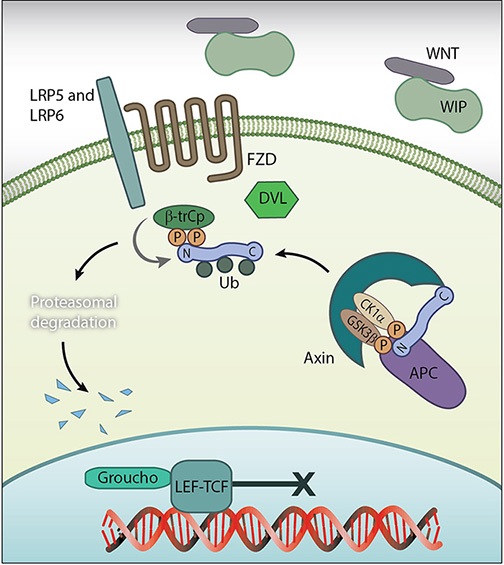
B-catenin in the Cytoplasm: Disassembly of the adhesion complex by proteases such as ADAM10 releases β-catenin into the cytoplasm. There, and in the absence of Wnt signalling proteins, it is subject to degradation by the β-catenin destruction complex. Figure adapted from [18432252]
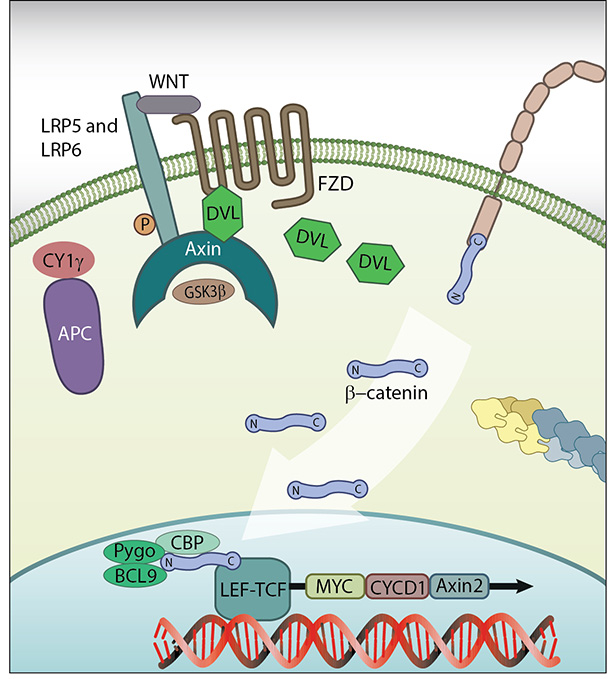
B-catenin in the nucleus: β-catenin in the nucleus: In the presence of Wnt signalling proteins, β-catenin enters the nucleus and interacts with the transcription factor LEF1 where it initiates transcription. Figure adapted from [11]
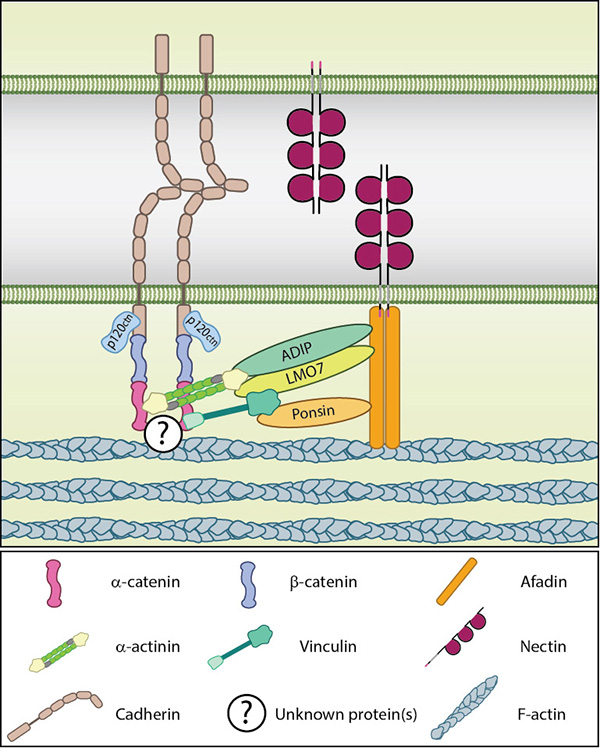
Basic components of the adherens junction.: Adapted from [18648374]. Nectin-based adhesions recruit cadherins to form the adherens junction. These receptors are indirectly linked through interactions at the cytoplasmic face. The proteins connecting the cadherin-catenin and nectin-afadin complexes are not known in their entirety, though several have been posited including; ponsin-vinculin, ADIP-α-actinin and LMO7-α-actinin. The latter complex is suggested to be involved in more mature adherens junctions and is therefore unlikely to be involved in the initial recruitment of cadherin by nectin [15140894]. The activities ofRho GTPases, such as Rac and Cdc42, also have a role to play in cadherin recruitment through their downstream effectors. These effectors include IQGAP1, which is recruited to early adhesion sites by nectins [12687012] and is suggested to aid in the recruitment of cadherin-catenin complexes to nectin-based adhesion sites by preventing the endocytosis of cell surface cadherins [18648374, 15263019].
Cadherin endocytosis upon release of cell-cell contact.: Eph4 mouse mammary epithelial cells plated on glass were treated with blebbistatin (an inhibitor of myosin II) in order to stimulate the release of cell-cell contacts. Cells were fixed 15 minutes later and stained for cadherin (red) and actin(green). Merge shown to the right. Cadherin can be seen accumulating in vesicle-like structures in the vicinity of the disassembling junction (inferred from the cortical actin denoting the cell edge). (Scale bar 2µm) [Image courtesy of Megha Vaman Rao and Ronen Zaidel-Bar, Mechanobiology Institute, Singapore].
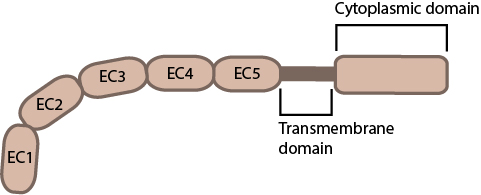
Cadherin extracellular domain structure: When bound to calcium the extracellular domain of cadherin adopts a curved conformation. EC1 (red), EC2 – EC5 (blue). (PDB file 1L3W from www.rscb.org)
Capping proteins promote actin filament disassembly: (A) Protrusion of filopodia or lamellipodia is dependent upon actin polymerization at the barbed end of actin filaments. (B) Capping proteins bind to existing actin filaments at the barbed end to prevent filament assembly.(C) ADP-actin continues to depolymerize from the pointed end of the actin filaments. When all of the actin filaments are capped in a filopodium, protrusion is prevented and filament disassembly is favored.
33.
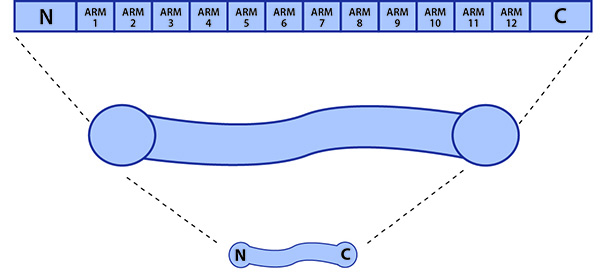
β-catenin: β-catenin contains three distinct functional domains, with the middle domain containing 12 ARM repeats.
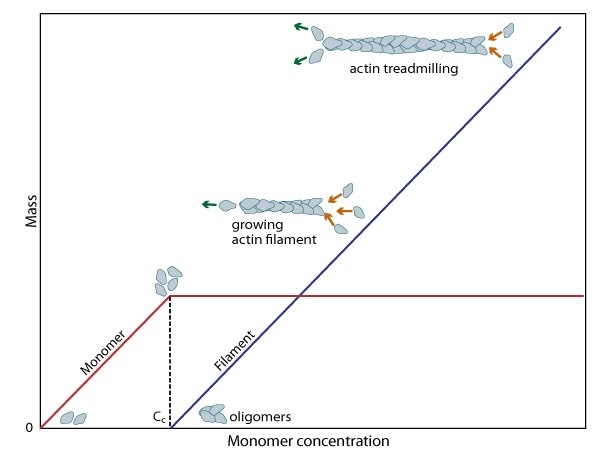
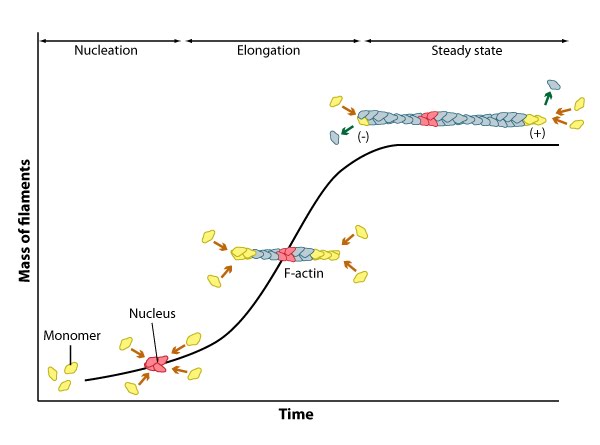
Cc and actin filament assembly.: The critical concentration (Cc) marks the level at which G-actin monomers are in equilibrium with the actin filaments. Actin filaments are only formed at monomer concentrations above the Cc
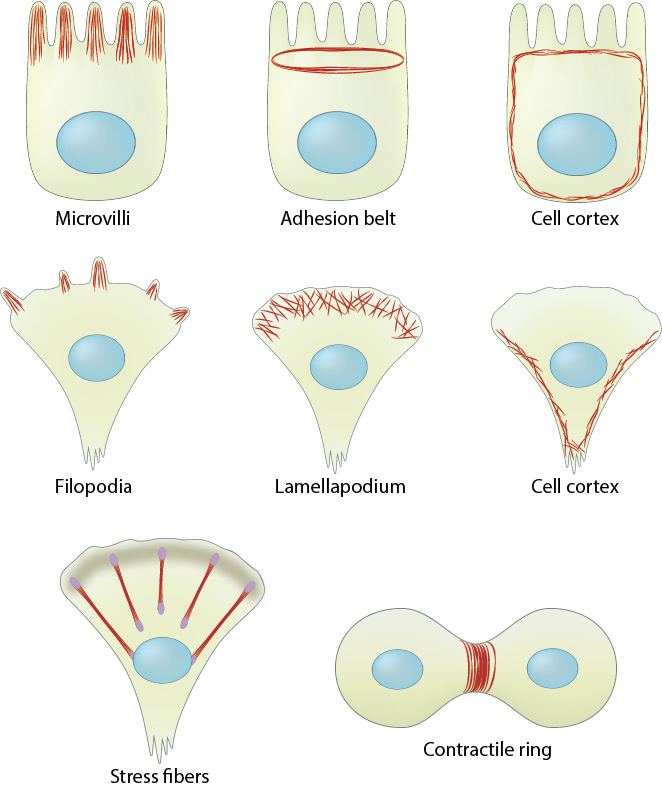
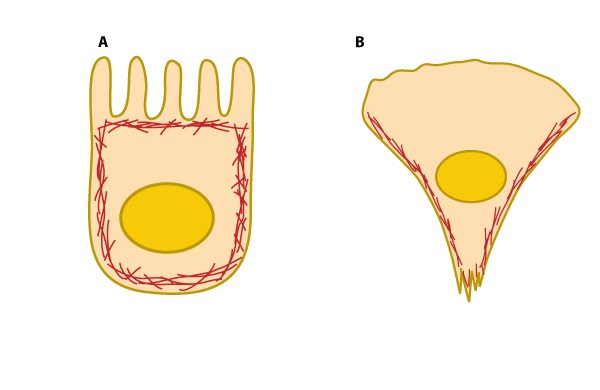
Cell cortex (aka cortical actin, actin cortex): (A) Cortical actin filaments (shown in red) are concentrated just beneath the plasma membrane in most cell types. (B) Migrating fibroblasts grown in 2D tissue culture have more cortical filaments on the dorsal (upper) surface than the ventral (lower) surface and they are concentrated towards the trailing edges [1]
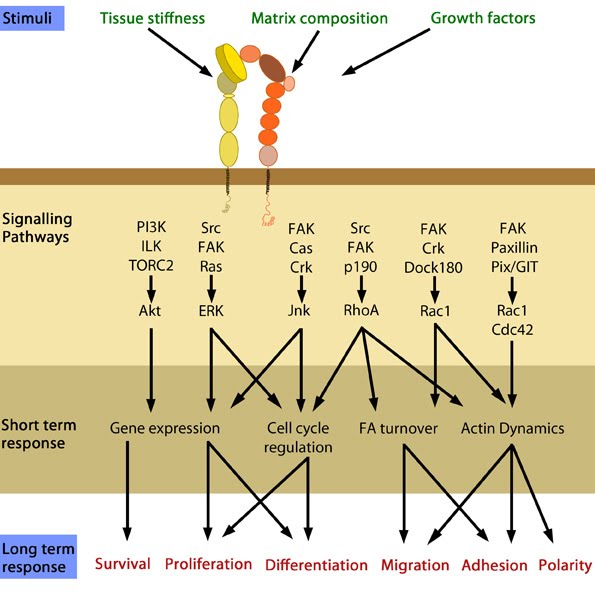
Cellular responses elicited by integrin signaling: In response to physical/chemical properties of the matrix and growth factors in the environment (outside-in signaling), integrins bind ligands and get activated. Accordingly, a variety of signaling pathways can be triggered mainly through the different kinases as mentioned above. These can bring about changes in one or more cellular events (short term responses) that eventually result in global (long term) responses in cellular behavior. Adapted from [37].
40.
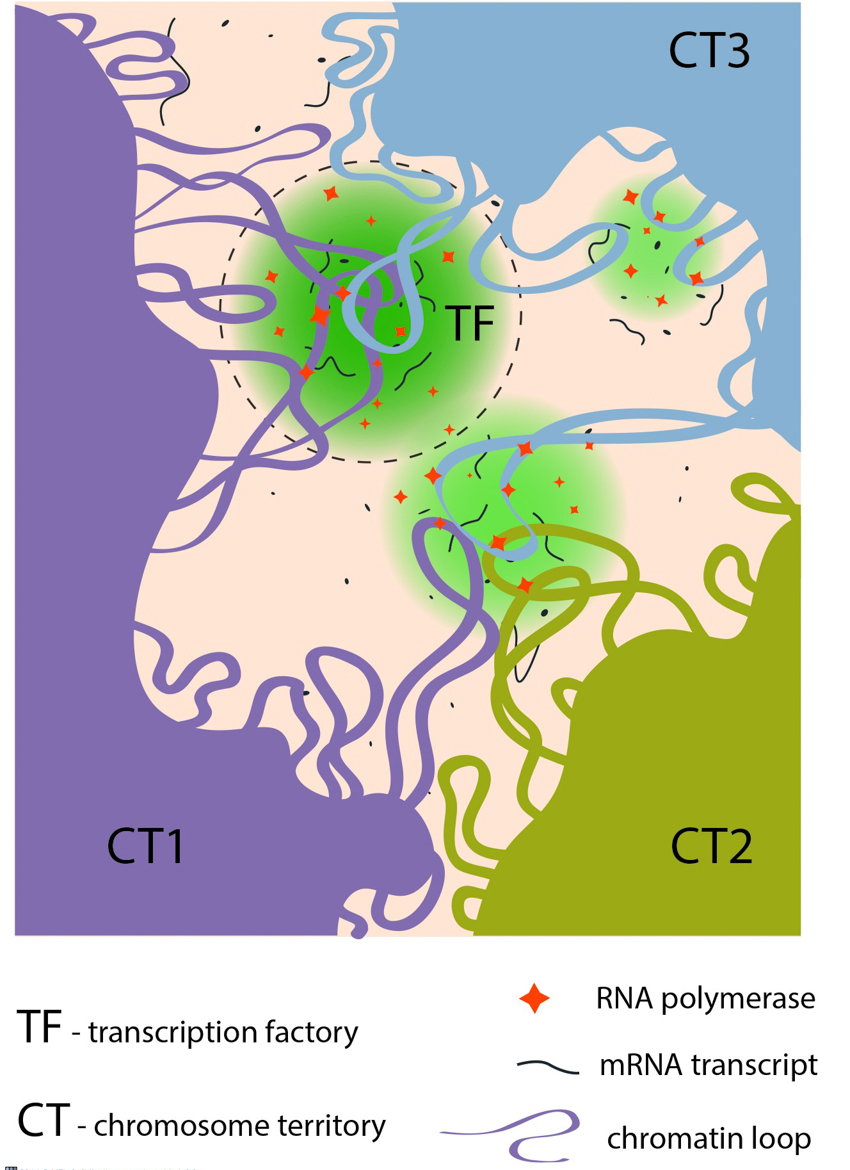
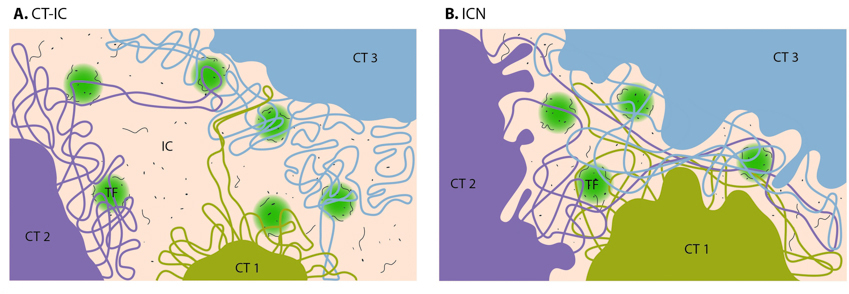
Chromatin loops share transcription factories: Chromatin loops from the same or from different chromosome territories often share transcription factories (TFs). TFs are enriched in RNA polymerase II complexes that produce nascent RNA transcripts, using co-transcribed genes from chromatin loops as their templates.
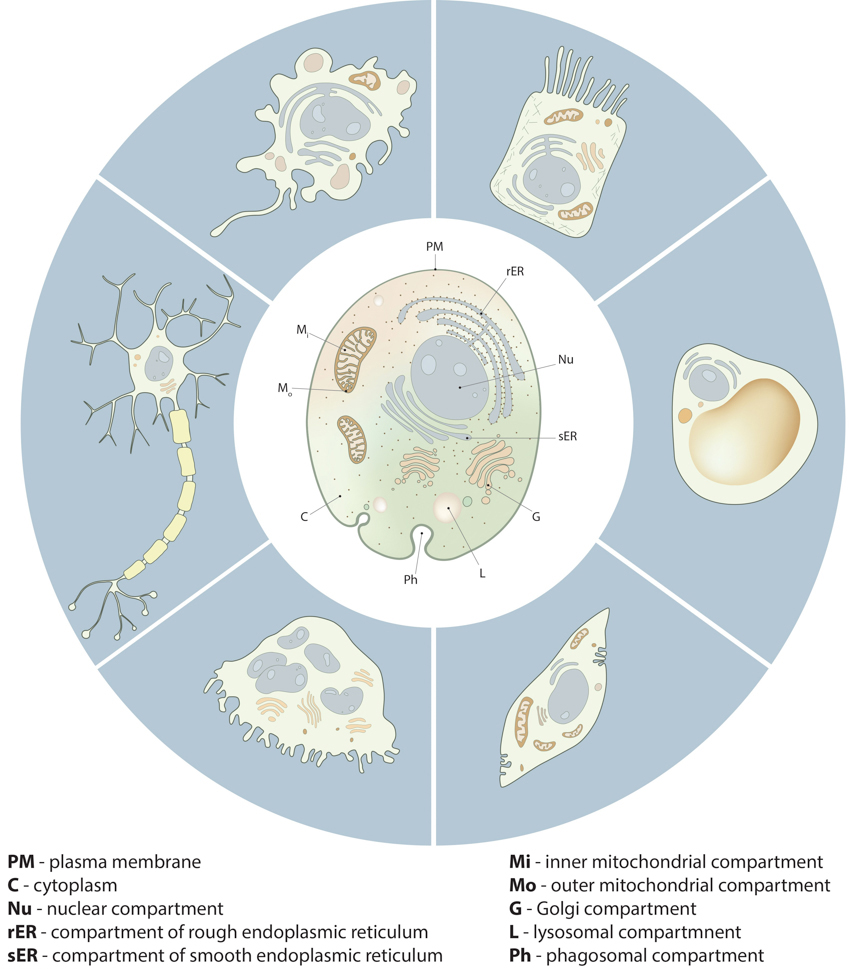
Compartmentalization in cells:: Despite the morphological and functional variety of cells from different tissue types and different organisms, all cells share important similarities in their compartmental organization. These fundamental compartments, often referred to as organelles, are summarized in the drawing of the generic animal cell (central cell). Examples of specialized cell types, shown around the generic cell, include neuron, macrophage, intestine epithelial cell, adipocyte, muscle cell and osteoclast.
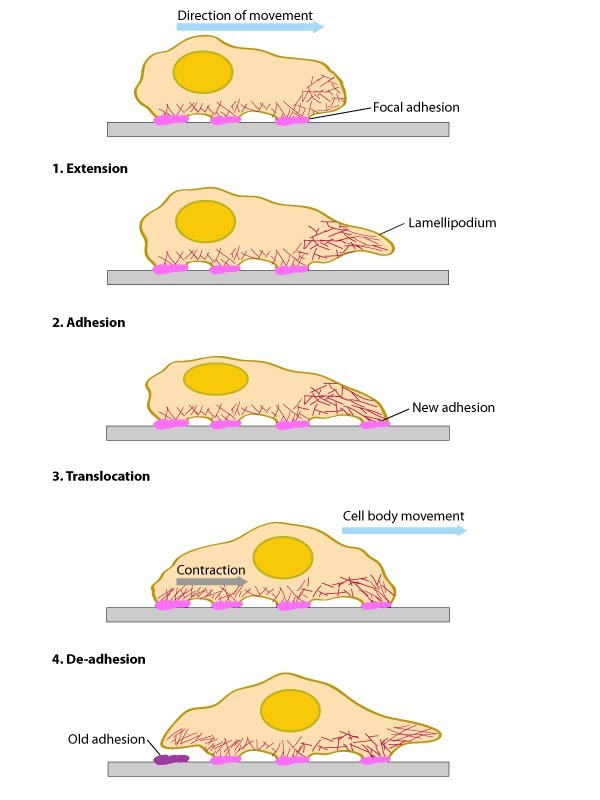
Conserved steps in cell spreading and movement: 1. Polymerization of actin filaments at the leading edge is translated into protrusive force. 2. Membrane protrusion facilitates the binding of transmembrane cell surface receptors to the substratum components. New adhesions are rapidly linked to the network of actin filaments. 3. The combined activity of retrograde actin movement and contractile forces produced by stress fibers generate tension to pull the cell body forward. 4. The forces produced by the contractile network combined with actin filament and focal adhesion disassembly, helps to retract the trailing cell edge. (Note: the "space" between the cell and the substrate [shown as a gray bar] is exaggerated in this diagram).
Contraction of transverse arcs: Contraction of the actin-myosin system allows the filaments to adopt a more regular structure that is contractile along its entire length; the extent of actin filament crosslinking may regulate these contractions. Myosin II motor protein activity is necessary for complete transverse arc formation (figure adapted from [16651381]).
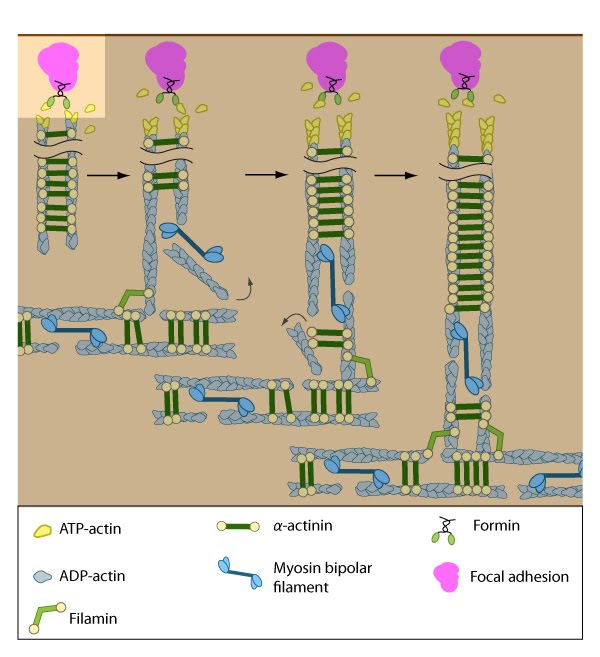
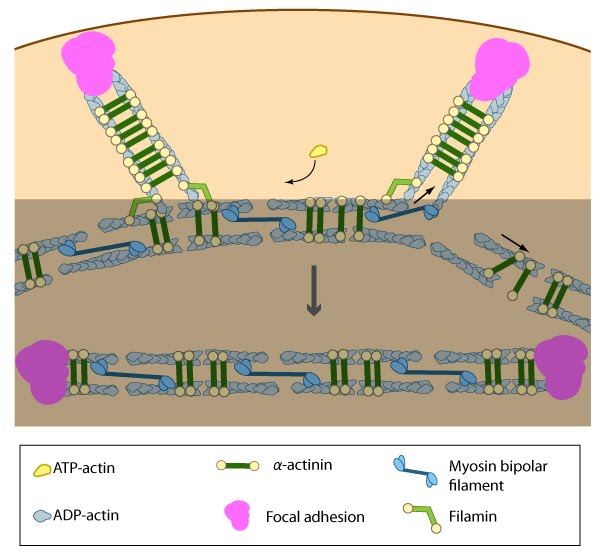
Contraction of ventral stress fibers: Once the actin filaments and myosin bundles are aligned, contractile activity along the filaments further organizes the bundles and brings the stress fiber to a completely ‘fused’ state: the completed ventral stress fiber is anchored by focal adhesions at both ends and the contractile bundles are dispersed throughout.
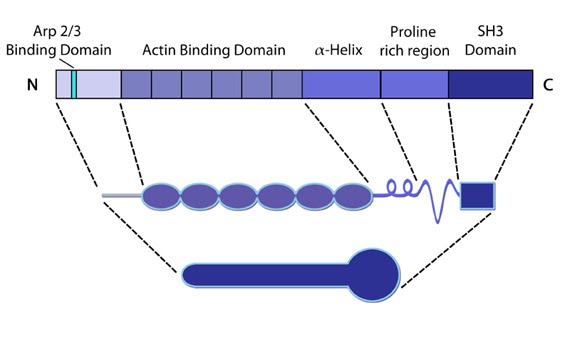
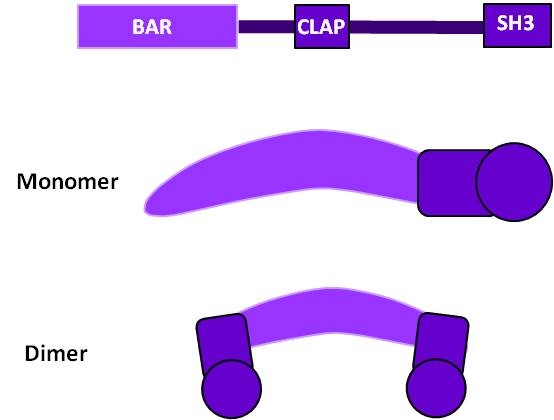
Cortactin: This schematic diagram illustrates the molecular organization of cortactin (reviewed in [2])
and provides examples for how cortactin is represented in figures
throughout this resource. Relevant domains believed to be important for
cortactin activity are highlighted.
49.
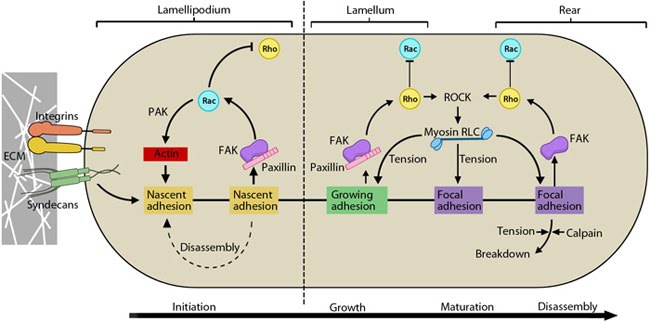
Cyclic activation of Rac and Rho regulate adhesion dynamics during migration: The schematic highlights the signaling pathways that play a significant role in mediating GTPase-regulated protrusion in the lamellipodia and the differential adhesion dynamics various regions of the cell. FAK signaling is important at all stages of the adhesion life cycle while kinases such as PAK and ROCK influence later stages by promoting actomyosin contractility. PAK is highlighted in green during maturation to indicate that is it activated earlier by Rac and not by Rho. Adapted from [17, 18].
50.
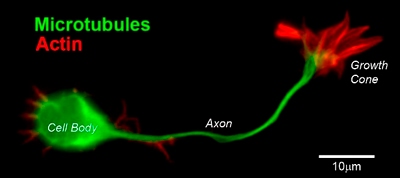
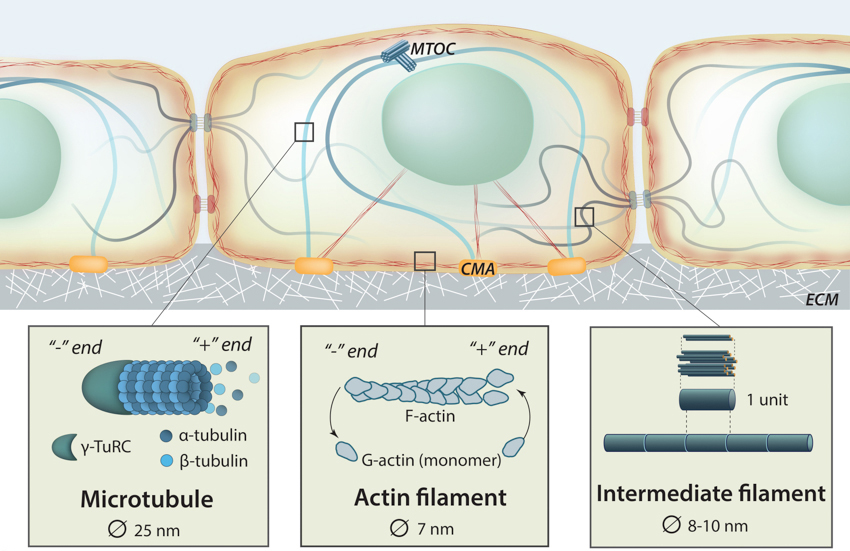
The Cytoskeleton: Three main components of the cytoskeleton include actin filaments (also called microfilaments), microtubules and intermediate filaments. They are distinct structural compositions that exhibit slightly different yet interdependent functions.
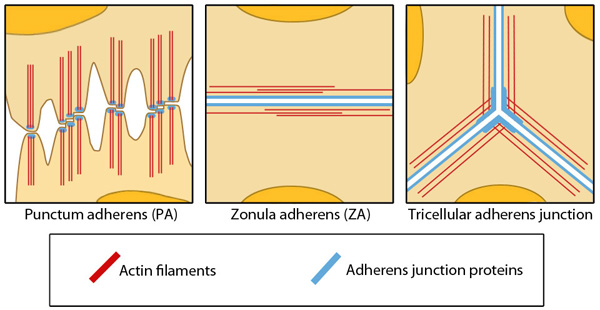
Different Structures of Adherens Junctions: Adapted from Yonemura S, 2011. This diagram depicts three different examples of adherens junctions; punctum adherens that are common in mesenchymal and neural cells, zonula adherens that are common in endothelial and epithelial cells and tricellular adherens junction, common in all cell monolayers [Franke WW et al, 2009].
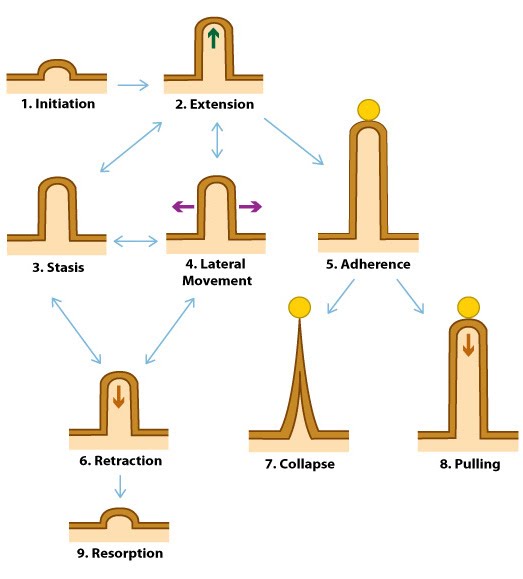
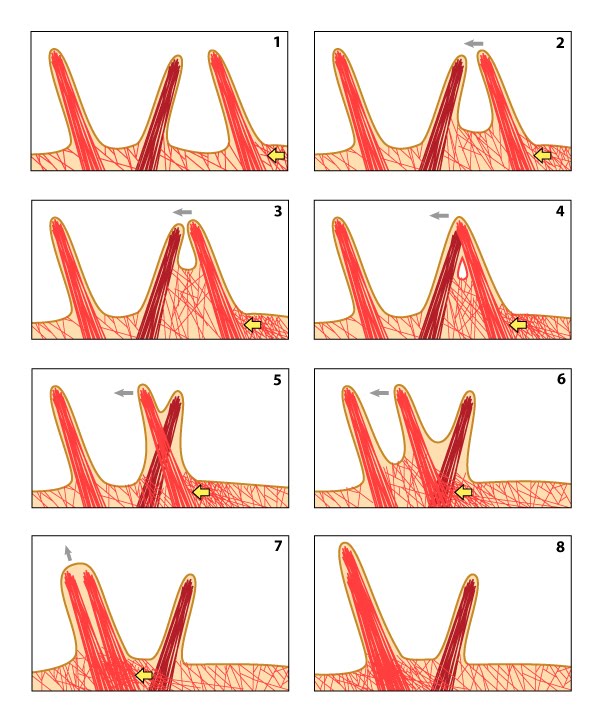
Dynamic behaviors of filopodia: Filopodia undergo 9 distinct steps in their formation. Double-sided arrows symbolize the ability of a filopodium to oscillate between different states.
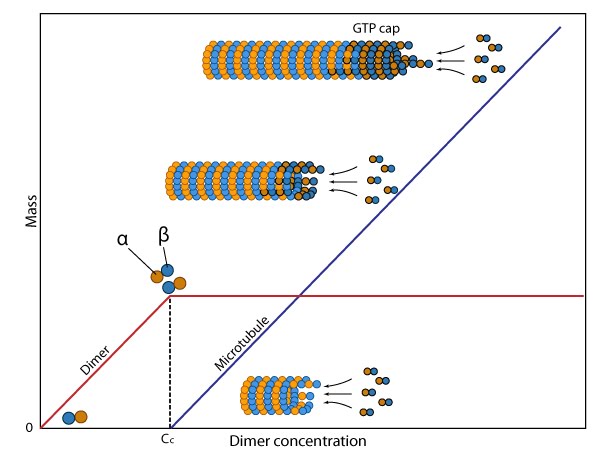
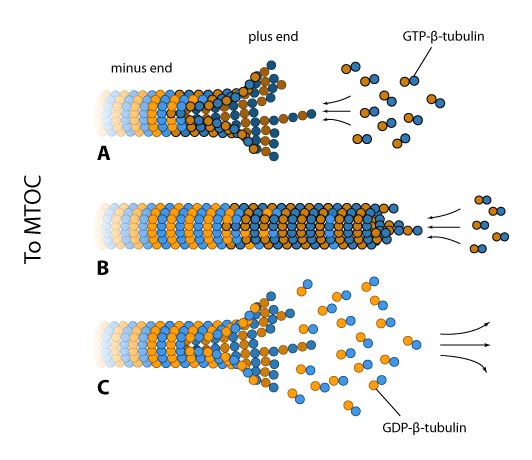
Dynamic instability of microtubules: (A) The intracellular concentration of tubulin found in most cells (10-20µM) favors microtubule assembly at the plus end. This end may appear frayed in slow growing filaments due to fewer lateral interactions between the protofilaments and because GDP-tubulin dimers have an inherent curve. (B) At high free GTP-tubulin dimer concentrations, hydrolysis is outpaced by rapid assembly at the plus end, thereby forming a rigid GTP-cap. (C) Concerted GTP hydrolysis at the plus end weakens the tubulin dimer interactions and the protofilaments rapidly disassemble.
58.
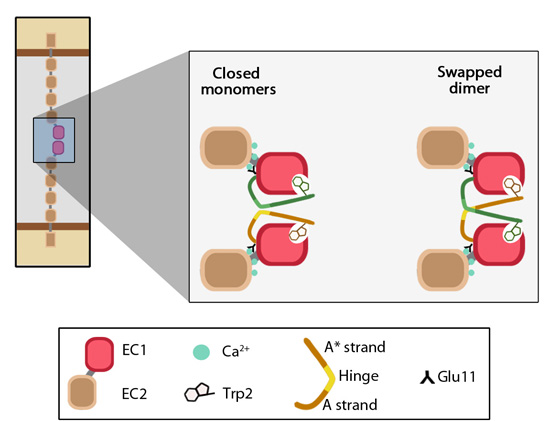
EC1-EC1 strand swapping.: Adapted from [21572446]. The outermost EC domain (EC1) of cadherins engage in trans interactions by forming a dimer interface. In the closed monomers of cadherin, β-strand A of the EC1 domain is held under conformational strain through anchoring via tryptophan-2 at one end and Ca2+-bound-glutamine-11 at the other end This conformation favors the ‘swapping’ of β-strands on apposing cadherin EC1 domains. In the swapped dimer, the side chain of the conserved tryptophan-2 of one EC1 domain to tucks into the hydrophobic pocket present on the opposing EC1 domain.
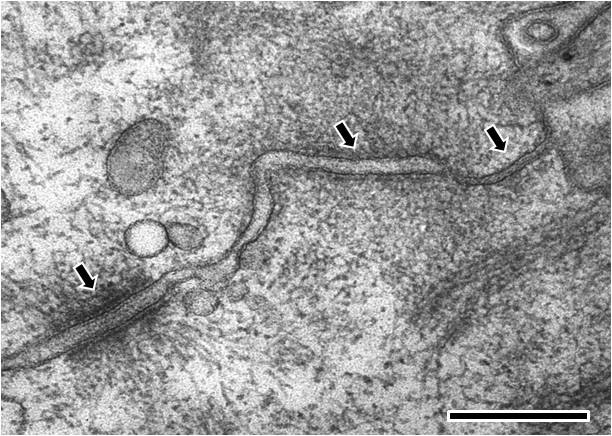
Electron microscopy image of rat intestinal mucosa epithelial cell-cell junctions: The cell is orientated such that the top right hand corner is apical and the bottom left hand corner is more basal.From left to right; the first arrow points to a desmosome with dense desmosomal plaques seen either side of the intercellular space of approximately 240 angstroms in width, the second arrow points to an adherens junction with an intercellular space of approximately 200 angstroms and the third arrow points towards a tight junction that shows close apposition of the plasma membrane leaflets, as inferred from the absence of an intercellular gap. (Scale bar 250 nm).
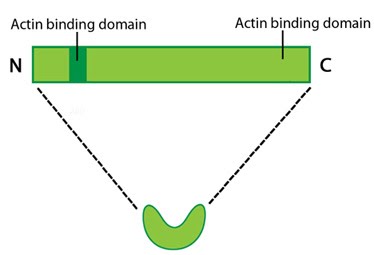
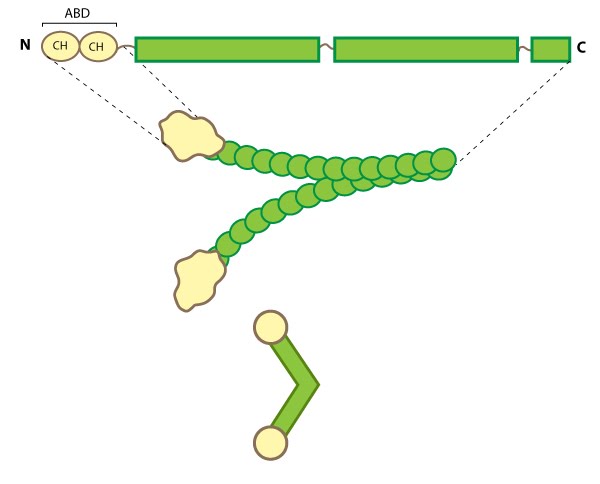
Filamin: This schematic diagram illustrates the molecular organization of filamin [2391361] and provides examples for how the filamin dimer is represented in figures throughout this resource. The ABD is at the amino (N)-terminus in contrast to the opposite end (the carboxy [C]-terminus), which contains a significant number of protein-protein interaction domains (reviewed in [15516996]).
65.
Filopodia can pull objects: After a filopodium binds to an object, retrograde actin movement and myosin motor activity provide the force(s) needed for pulling the object towards the cell body. Once pulling starts, the initial slow movement (#1) is followed by a burst of rapid movement (#2) that diminishes as the object reaches the cell body (#3) [2].
Fimbrin: This
schematic diagram illustrates the molecular organization of fimbrin and
highlights the relevant domains for binding to actin filaments.
67.
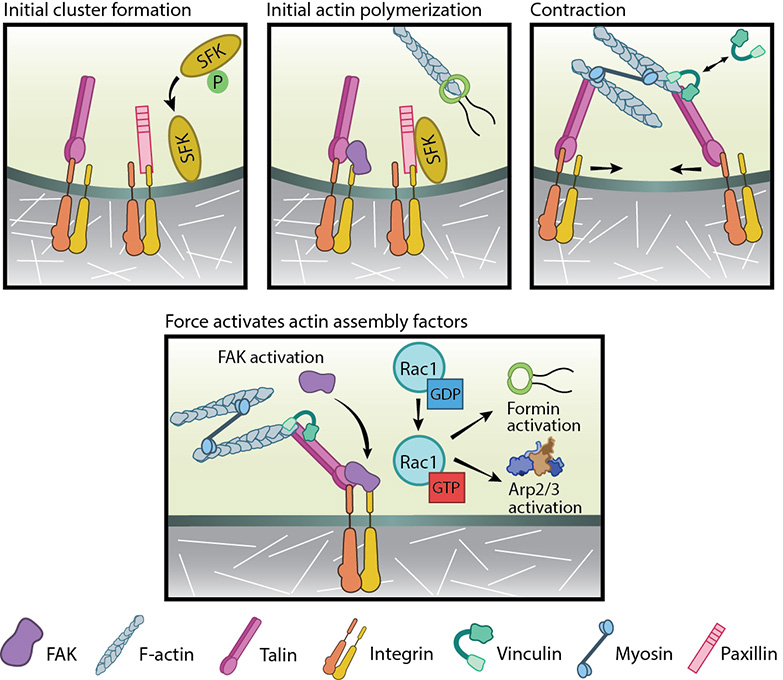
Focal adhesion initiation events: Adapted from [32]. Initial integrin clusters (top left), after activation by talin binding, provide avenue for initial actin polymerization (top middle) by recruiting focal adhesion components- FAK, SFKs and paxillin. New actin filaments tether to talin, the clusters get pushed away and then pulled closer by myosin contractions (top right). This causes cycles of transient talin stretching and vinculin binding until the talin- actin bond stabilizes. Upon stable vinculin binding (bottom), further integrin clustering and signaling promote Rac1 activation. Rac1, in turn, further activates actin polymerization modules, Arp2/3 and formins.
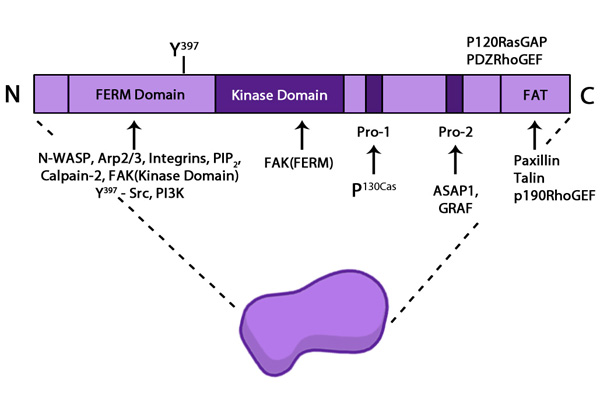
Focal adhesion kinase (FAK): The top
schematic diagram illustrates the
molecular organization of FAK and the bottom shape shows how it is
represented throughout this
resource.
69.
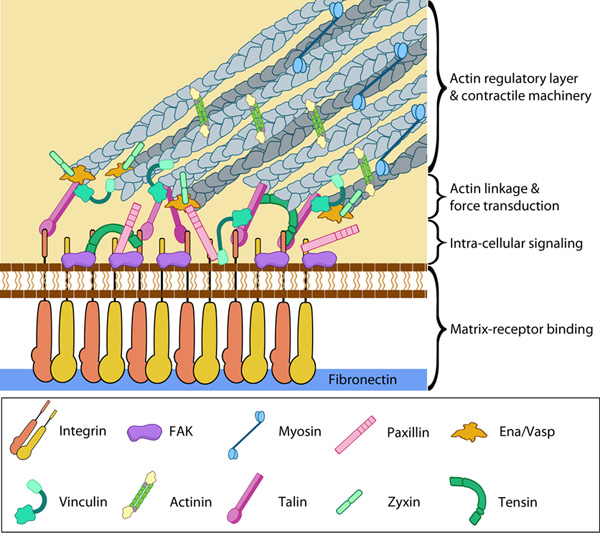
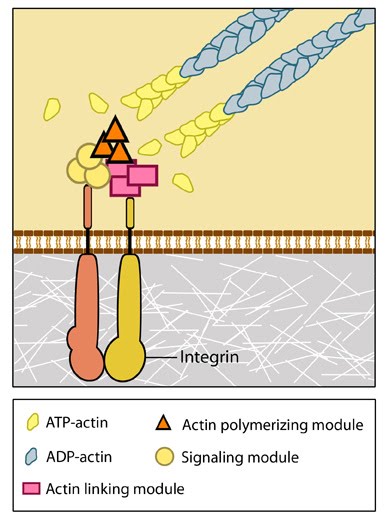
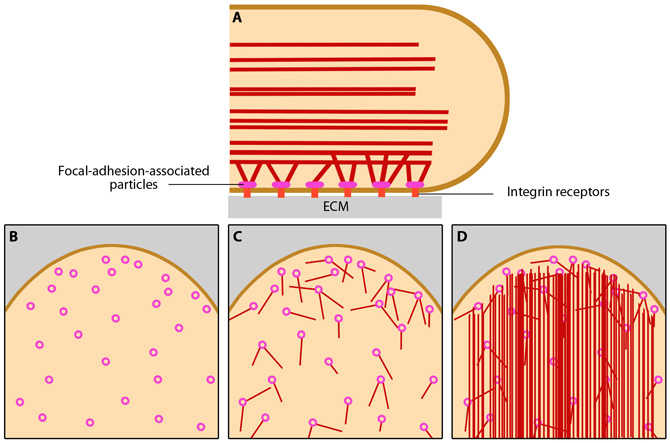
Focal Adhesion organization: A mature FA contains hundreds of proteins that are grouped based on their contribution to four basic processes: receptor/matrix binding, linkage to actin cytoskeleton, intracellular signal transduction, and actin polymerization. Both actin polymerization and actomyosin contractile machinery generate forces that affect mechanosensitive proteins in the actin linking module, the receptor module (e.g. integrins), the signaling module, and the actin polymerization module. The combined activity of the mechanosensitive components form the mechanoresponsive network. The theoretical organization and protein-protein interactions as shown are based upon references provided in the glossary pages for each protein. Relative position of the components is based on [35]
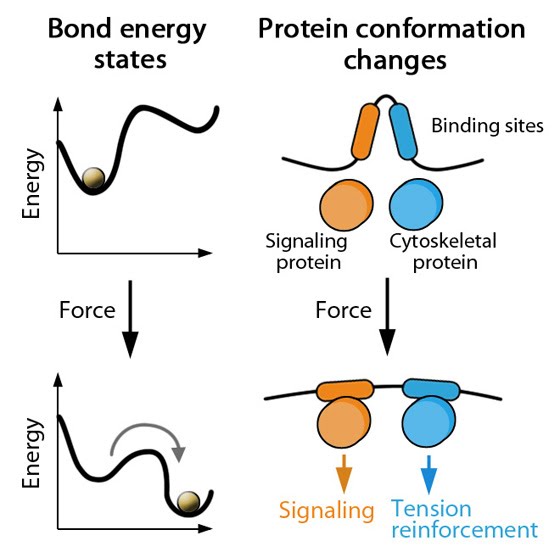
Forces can bring molecules together: When sources such as contractile stress fibers generate internal cell force(s), the force is transmitted throughout the array of actin filaments (shown in red). Mechanosensors, proteins, or complexes; (shown in green) that are linked to the cytoskeleton network can be locally concentrated. Thus, force not only causes direct deformation of mechanosensors to modify their activity, but clustering may also promote mechanotranduction by modulating enzyme kinetics, enzyme and substrate binding, etc. The cell membrane may also distort at sites where the cytoskeleton is linked to the plasma membrane, thereby influencing mechanosensors at the cell surface.
72.
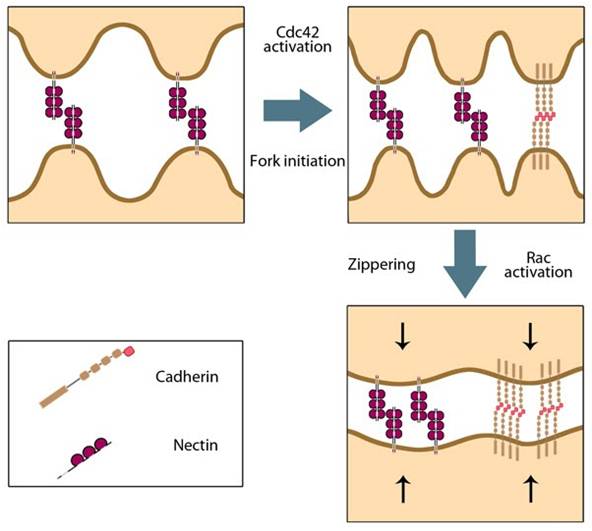
‘Fork initiation and zipper’ model for adherens junction formation: Initial cell-cell contacts are mediated by nectin-nectin trans interactions that activate the Rho GTPase Cdc42. Cdc42 stimulates an increase in filopodial protrusions during a phase termed ‘fork initiation’. This promotes more trans interactions between nectins as well as cadherins. Further downstream signaling via Cdc42 activates Rac GTPase and leads to the ‘zipper’ phase, whereby lamellipodial protrusions seal the gaps between filopodial cell-cell contacts.
Formation of ventral stress fibers: Actin filaments in ventral stress fibers are originally created either de novo or by extending pre-existing filaments. The filaments are anchored to focal adhesions and crosslinked into bundles by components of the actin linking module (e.g., α-actinin, filamin).
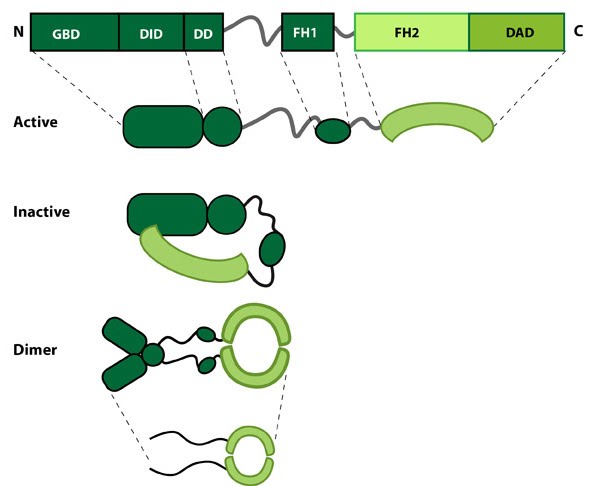
Formin: Formin.This schematic diagram illustrates the molecular organization of formin. Relevant domains/regions that are believed to be important for actin binding and protein-protein interactions are highlighted (reviewed in [2]. An intramolecular interaction between the diaphanous inhibitory domain (DID)and the diaphanous auto-regulatory domain (DAD), which prevents formin (e.g. mDia1) from nucleating actin filaments, is relieved by Rho binding to the GTPase binding domain (GBD, aka CRIB domain) [3]. However, this regulation may be more complex [4, 5]. DD = dimerization domain, FH = formin homology domain
75.
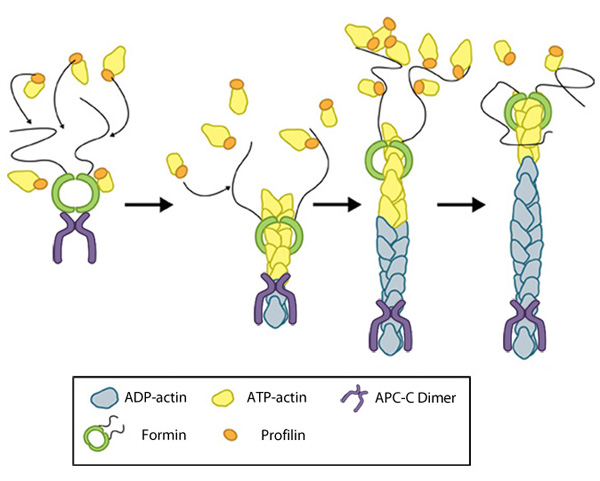
Formin-mediated nucleation of actin filaments: The FH2 domains of the formin dimer (shown in green)
bind to actin monomers to initiate filament assembly. Recent studies
indicate this is assisted, or even mediated, by additional factors such
as APC. The FH1 domains
of the formin dimer (shown as black lines) have short polyproline
sequences that interact with profilin. Profilin binds to both formin and
actin monomers to increase the addition of actin monomers to the barbed
end of the filament.
76.
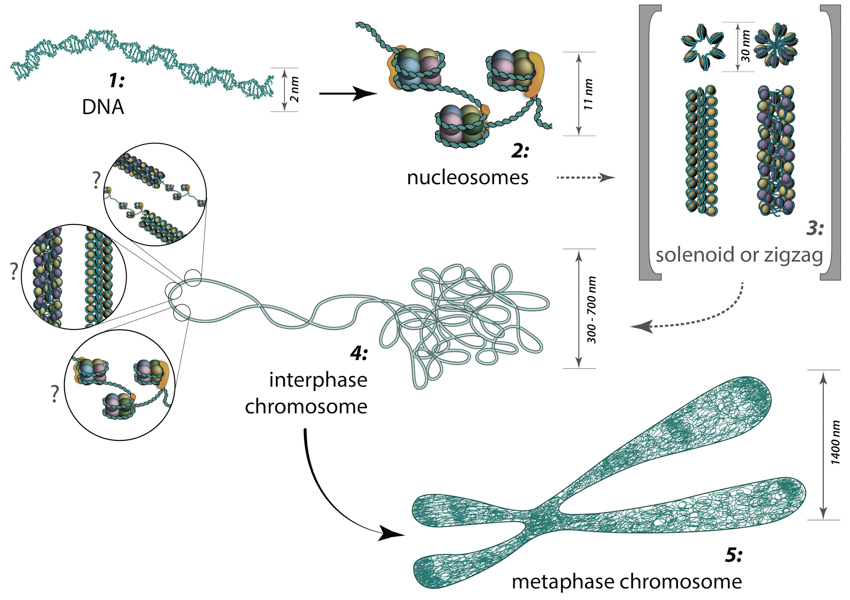
From DNA to metaphase chromosome: Folding of DNA into nucleosomes achieves initial 6-fold compaction level. Histone variants present in the nucleosome core, posttranslational modifications and linker histone H1 position can all control DNA accessibility for transcription at this compaction level. Further chromatin condensation into 30 nm fibers (i.e., zigzag or solenoid) is suggested by in vitro data and is yet to be proved or discredited to exist in vivo. During the interphase, chromatin is folded into 300-700 nm domains, which together comprise a chromosome territory. The structure and organization of chromatin loops inside a chromosome territory remains the matter of debates and was proposed to exist in the form of solenoid, or zigzag, or nucleosomes, or a hybrid of those.
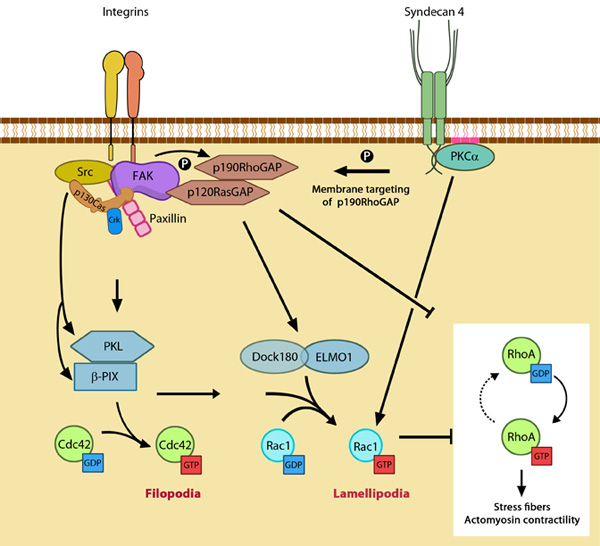
GTPase signaling at the leading edge mediated by integrin
β1 and syndecan-4: Integrin signaling and nascent adhesion formation recruits GEFs such as Dock 180 and PIX for Rac and Cdc42 at the leading edge, activating them. It also inactivates p190RhoGAP while PKC α activation downstream of syndecan-4 sequesters it the membrane to suppress Rho activation. PIP2 is indicated on the membrane in pink. PKCα activation also aids further Rac1 activation and ensures sustenance of Rac1 activity at the leading edge by lipid distributions (not shown). Adapted from [18].
81.
Incorporation of dorsal stress fibers: Dorsal stress fibers (DSF) appear to contact transverse arcs at their proximal ends towards the cell body. DSF are thought to 'feed' the transverse arcs with filaments of mixed polarity. Although myosin II bundles are generally absent from DSF, occasionally they will displace α-actinin and become incorporated towards the proximal end of DSF (figure adapted from [1869506]).
Initiation of dorsal stress fibers: Rho GTPases promote signal transduction cascades and interact directly with components of the actin polymerizing module to initiate actin filament assembly. Actin filaments in dorsal stress fibers are primarily initiated by formin-mediated nucleation (e.g., mDia1) (figure adapted from [16651381]).
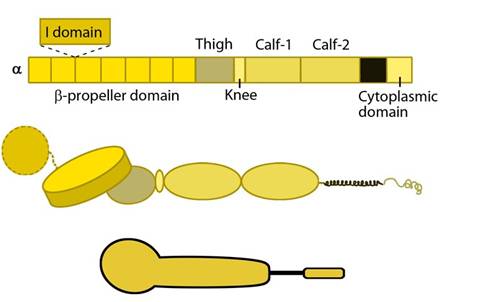
Integrin alpha chain: Integrin α subunit domains: Top: Linear domain arrangement. Middle: The globular structure formed by protein domains. Bottom: simplified version of the integrin α subunit. The αI domain is present in some subtypes of the α subunit.
85.
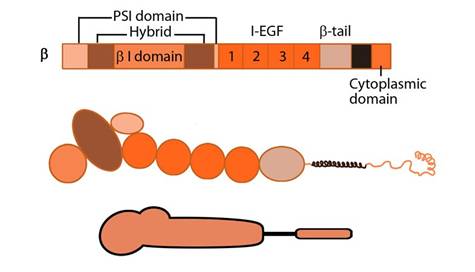
Integrin beta chain: Integrin β subunit domains: Top: Linear domain arrangement. Middle: The globular structure formed by protein domains. Bottom: simplified version of the integrin β subunit.
86.
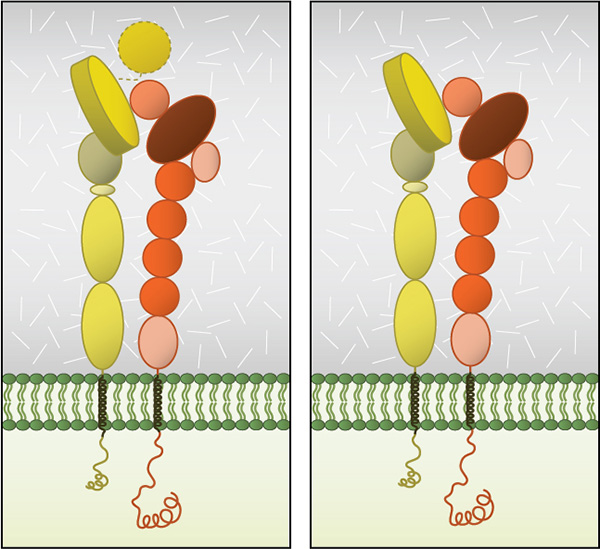
Integrin dimer structure: Globular domain structures of α and β subunits in a stable dimer. Ligand binding happens at the interface of the αI (left panel) or β-propeller (right panel) and the βI domain.
87.
Integrins as adhesion receptor in focal adhesion (FA): Integrins upon binding to ECM ligand gets activated and undergoes conformational changes. This leads to a series of events including i) recruitment of binding proteins to the site leading to actin linking, ii) activation of signaling molecules leading to integrin clustering and iii) actomyosin contractions leading to adhesion strengthening. All these result in iv) formation of new actin filaments by actin polymerization modules.Cell-matrix adhesion receptors are grouped according to the ligand that is bound. In some cases, adhesion receptors bind more than one type of ligand (e.g. the integrin family) and different receptor groups may cooperate with each other to bind their ligands (reviewed in [18583124, 19355970]).
88.
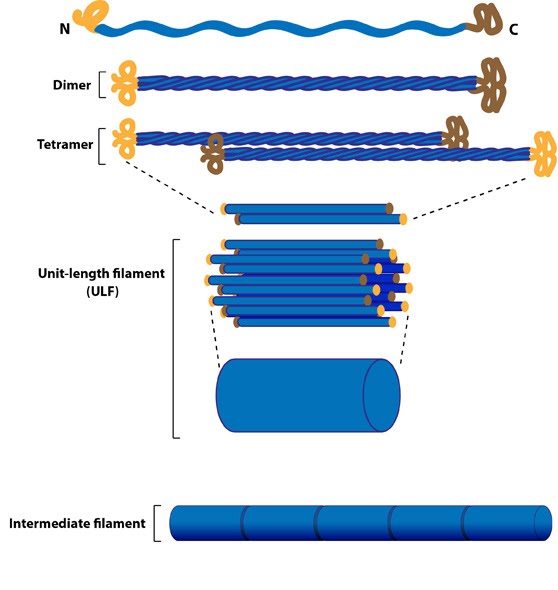
Intermediate Filament Assembly: Intermediate
filaments are built from monomers that associate with each other form
dimers. Pairs of dimers then associate in an anti-parallel fashion to
form staggered tetramers. Lateral associations between eight tetramers
form unit-length filaments, which are able to anneal to each other,
end-to-end, to form intermediate filaments.
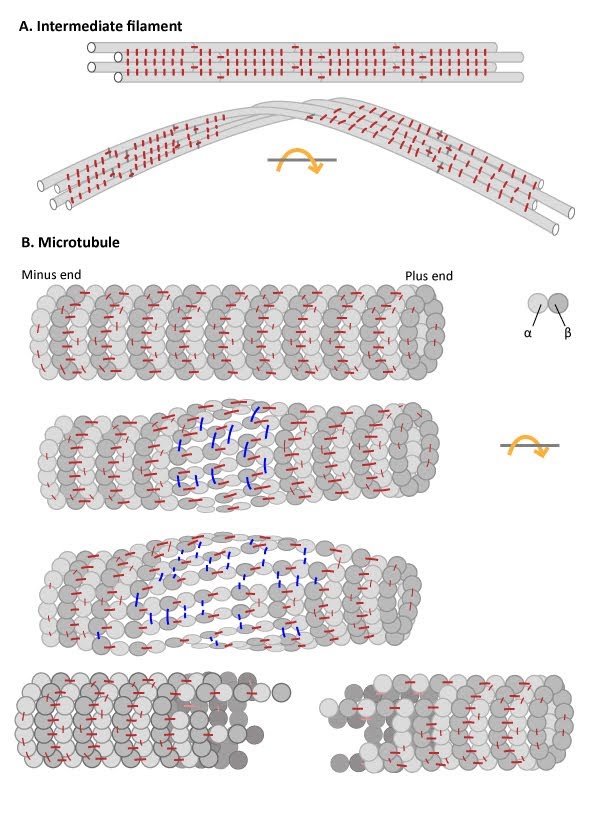
Intermediate Filaments are flexible and resistant to force: (A) Numerous
lateral interactions and fewer longitudinal interactions between
constituent protofilaments within an IF generates high tensile strength
and makes IFs resistant to compression, bending, twisting and stretching
forces. (B) Microtubules have more longitudinal interactions
between constituent tubulin dimers within protofilaments and fewer
lateral interactions between protofilaments. The stable longitudinal
interactions within individual protofilaments promote rigidity within
microtubules, making them resistant to bending and compression forces.
In contrast, the weaker lateral interactions between protofilaments are
susceptible to breakage when stressed by twisting forces.
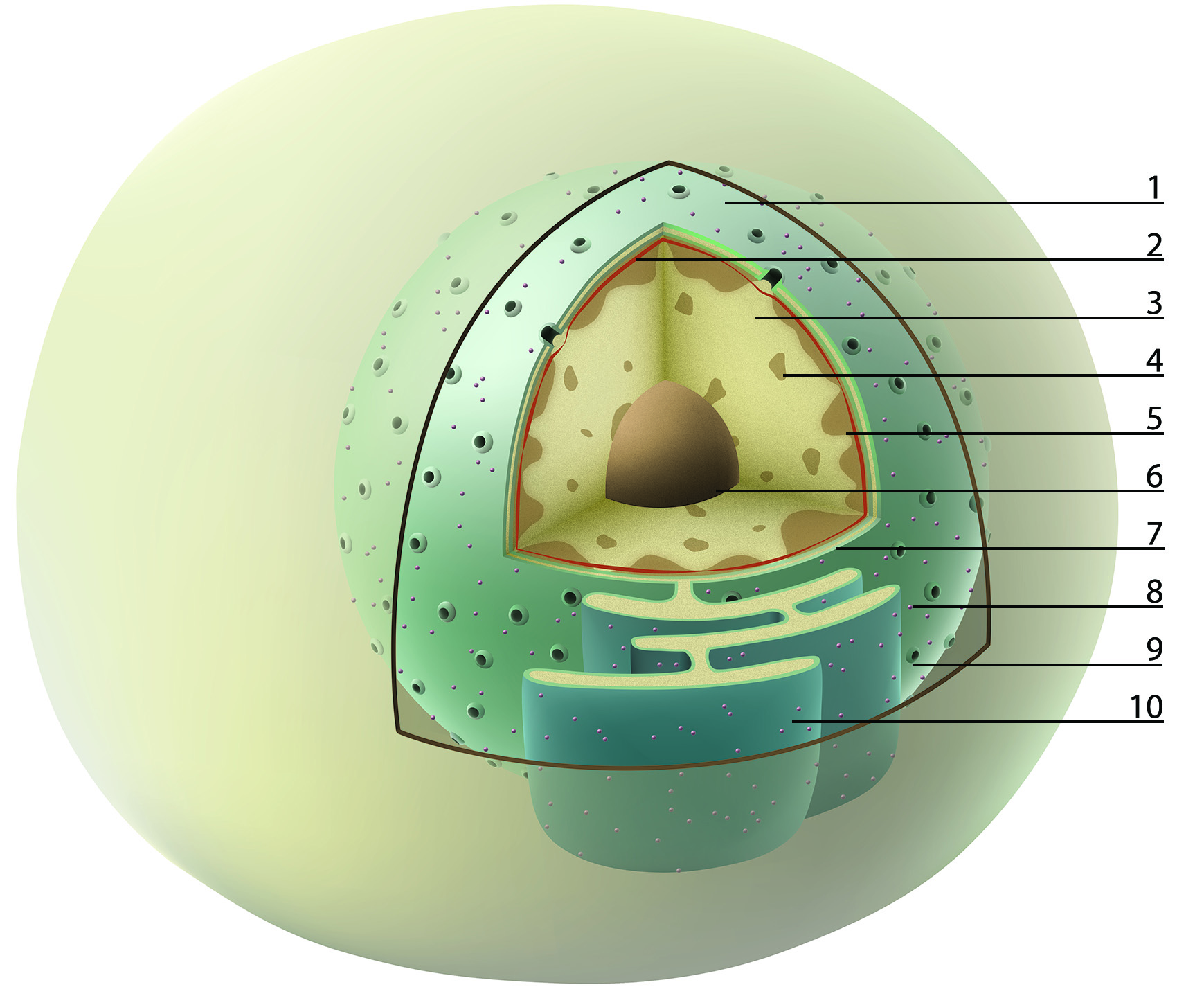
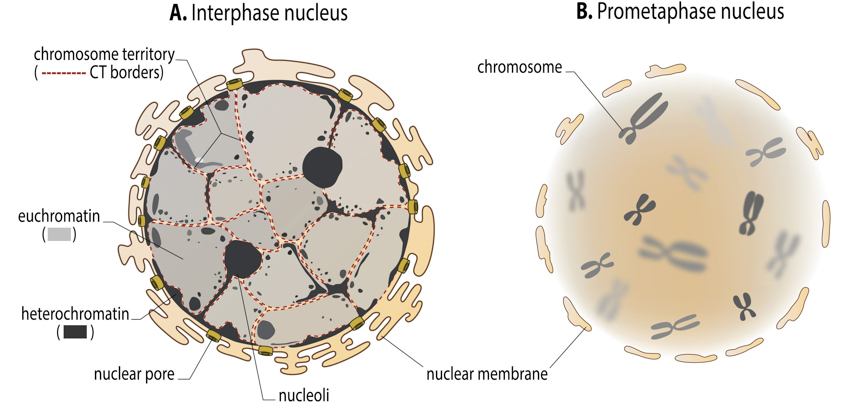
Interphase and mitotic chromatin: In the interphase nucleus, chromosomes are difficult to distinguish from each other. Never the less, they do occupy a discrete space inside a nucleus – so called chromosome territory (borders of chromosomes territories are suggested as red dotted lines in the figure A). Lighter stained euchromatin (transcriptionally active) and the patches of darker heterochromatin (transcriptionally silent) are, on the other hand, easy to visualize. During the cell division, chromosome territories transform into highly condensed chromosomes, which then can be clearly distinguished from one another. Together, mitotic chromosomes, visualized in light microscope, are called karyotype.
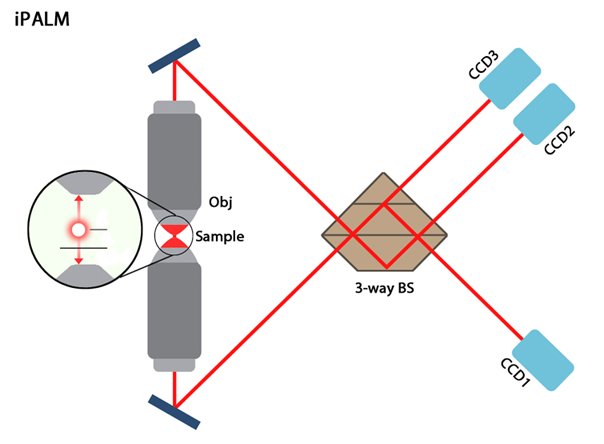
iPALM microscope schematic.: Image adapted from [7]. The distance emitted photons will travel in the two paths differs depending on the flurophores axial position (and hence the distance from each objective lens). The two signals are recombined using a 3-way beam splitter (BS) which causes self-interference of the photons and produces three beams of varying intensities which are subsequently detected by CCD cameras.
IRSp53: This schematic diagram illustrates the molecular organization of IRSp53
and provides examples for how IRSp53 is represented
in figures throughout this resource.
93.
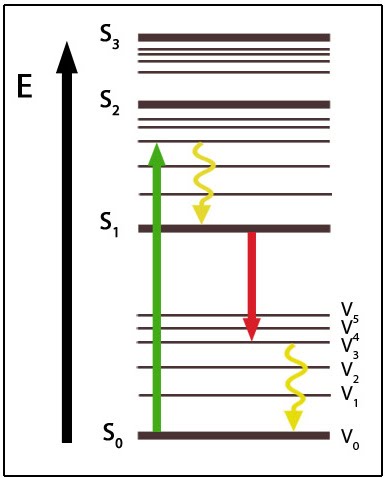
Jablonski Diagram of Energy States: This diagram shows various energy states (S) of fluorescence. Following excitation to a higher energy state (green) the fluorophore will undergo vibrational relaxation (yellow) before emitting fluorescence at a longer wavelength (red) and undergoing vibrational relaxation back to the ground state (S0V0).
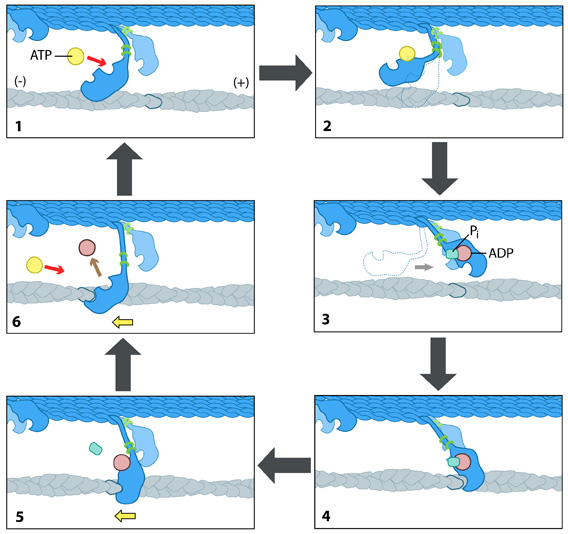
The kinesin powerstroke: In the initial stage of the powerstroke cycle, the neck linker of the leading head is tethered to the head but extended backwards and not interacting with the docking site (A) Following ATP binding and the subsequent conformational change, the neck linker docks with its docking site in the leading head. This pulls the trailing head forward (B). The trailing head continues forward until the neck-linker of the trailing head disengages the docking site. This head finds the next binding site and becomes the leading head (C). The cycle continues, allowing the Kinesin to "walk" down the microtubule filament.
96.
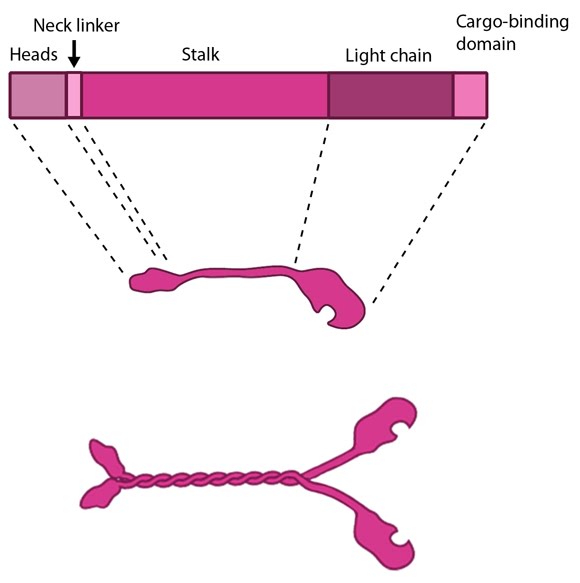
Kinesin schematic: The kinesin protein contains a head domain that binds to microtubules, a stalk region where two kinesin monomers interact and dimerize, and a cargo-binding domain that allows the protein to facilitate cytoskeleton-dependent intracellular transport.
97.
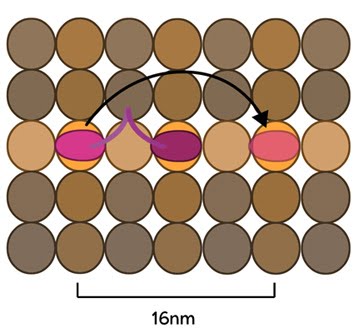
Kinesin stepping schematic: The kinesin head domain binds to microtubules. Each step involves a translocation of the trailing head to a binding site 16nm ahead of the previous site. It is then considered the leading head.
98.
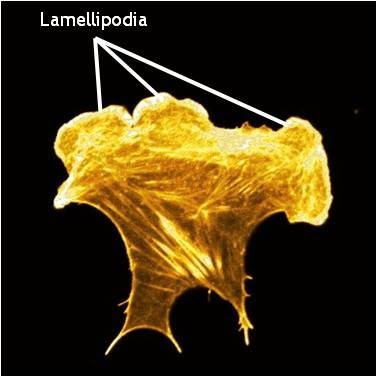
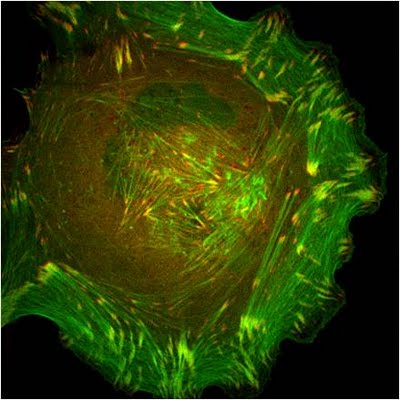
Lamellipodia in a cell stained for F-actin: A mouse embryonic fibroblast of the RPTPa cell line, plated on a fibronectin coated glass cover slip. The cell was transfected with RFP-Lifeact (a kind gift from Dr Roland Wedlich-Soldner, Max Planck Institute of Biochemistry, Germany), which labels F-actin in living cells. The cell was imaged using a Nikon A1Rsi confocal microscope at 60x magnification and false coloured yellow. [Image captured by Wei Wei Luo, Mechanobiology Institute, Singapore]
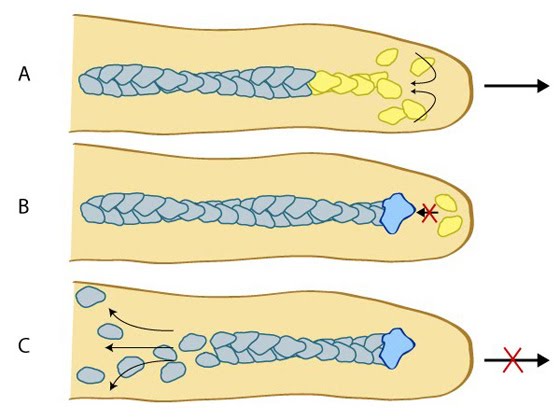
Lateral movement of filopodia: The direction of cell movement in a migrating cell is primarily controlled by actin filament assembly at the leading edge. In the diagram above, increased actin polymerization at the side of the filopodium (yellow arrow) pushes the membrane forward and to the left (panel #1). The forces produced by actin polymerization against the actin bundles of a filopodium can lead to lateral movement (panels #2 & 3). Under the microscope, filopodia appear to cross-over one another as distinct units when they are separated within the three-dimensional framework of the cell (panels #4-6). When a laterally moving filopodium encounters another filopodium within the same three-dimensional space, the two filopodium can fuse to become one; this frequently increases the width and length of the resulting filopodium (panels #7 & 8).
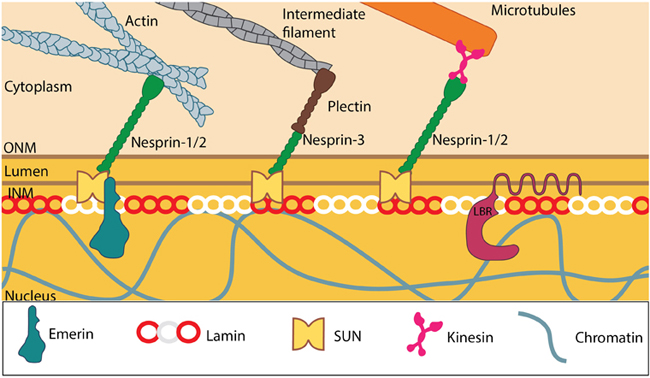
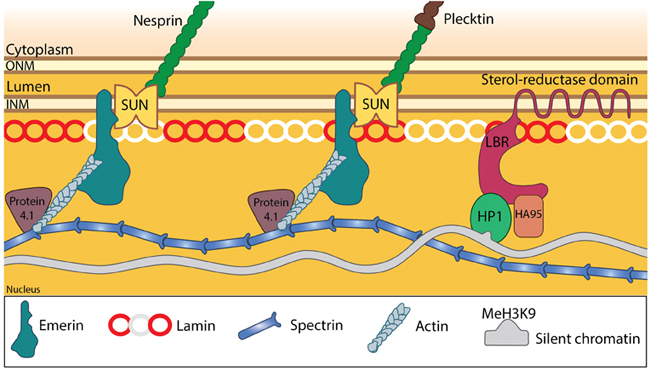
LINC complexes provide structural support to the nucleus: SUN and KASH domain proteins (mainly nesprins) provide the essential linkage between the three main cytoskeletal networks aend the nucleokeleton especially the nuclear lamina. Plectin acts as an intermediary linker connecting the intermediate filaments while a motor protein (e.g: kinesin) is generally involved in linking microtubules. INM- Inner nuclear membrane; ONM- Outer nuclear membrane; LBR- Lamin B receptor. Together these provide the outward pull that maintains the nucleus in a stretched state. Adapted from [21321324, 15688064, 21327104].
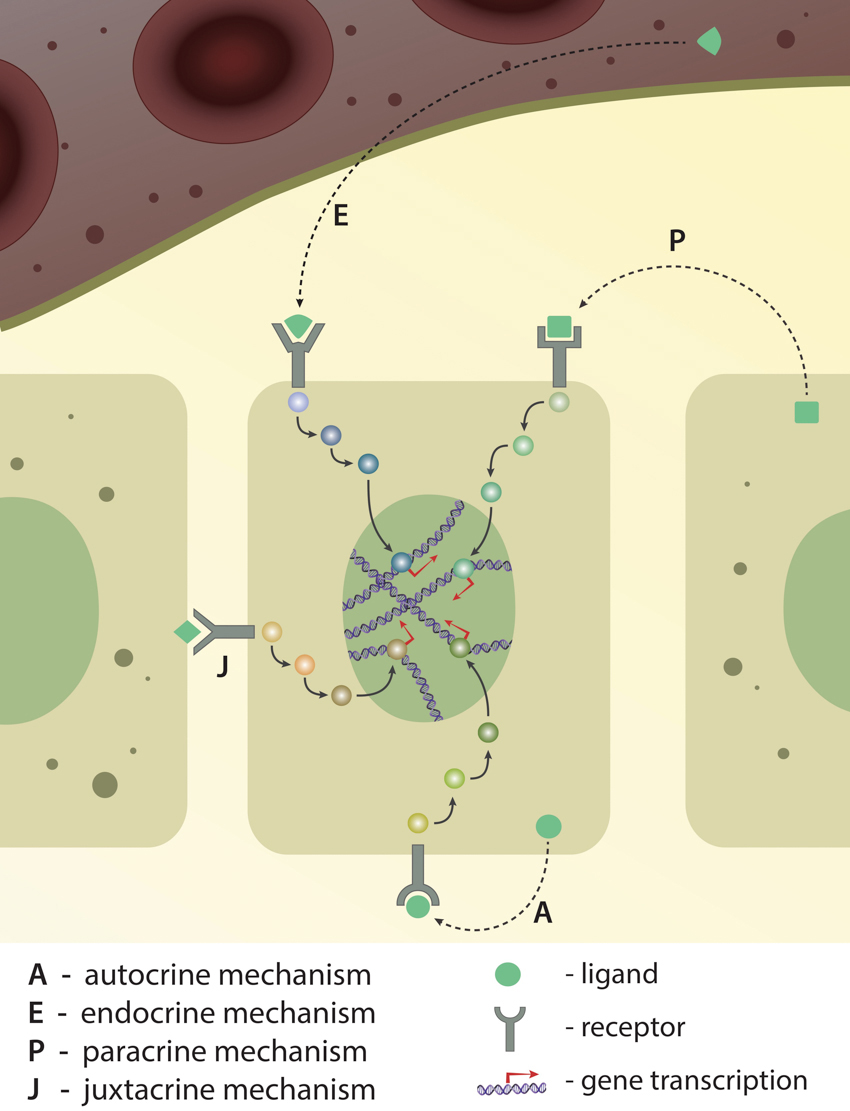
Long-distance versus local communication: Depending on the ligand's origin (from the same cell, from the neighbour cell or from far distance), recptor-ligand interaction and signaling pathway activation is classified into four different types: autocrine, endocrine, paracrine and juxtacrine. Activation of either of these pathways ultimately results in gene expression.
Matrix metalloproteinases (MMPs): This schematic diagram illustrates the molecular organization of MMPs (reviewed in [3]). Relevant domains believed to be important for MMP activity are highlighted (reviewed in [3]).
103.
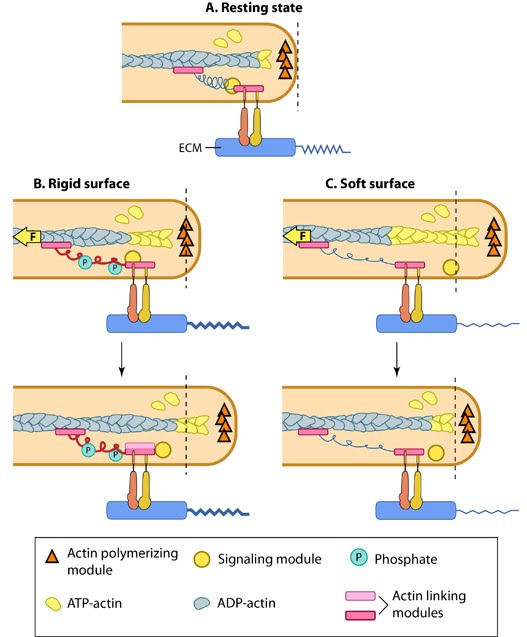
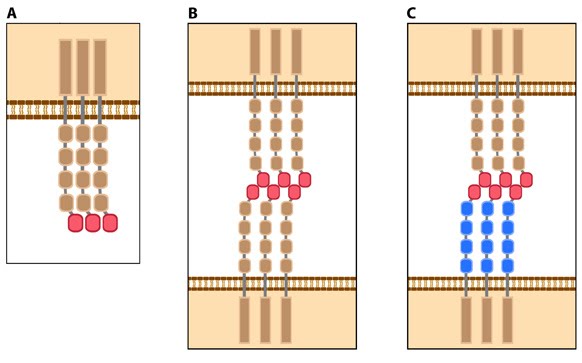
Matrix property affects FA dynamics and mechanotransduction: This figure illustrates how force (e.g. substrate rigidity, contraction) hypothetically alters the position of molecules and influences subsequent mechanotransduction events (adapted from [16, 17]). A. In the resting state, actin filaments are linked to the ECM through flexible protein linkers (shown as dark pink ovals with a stretchable coil domain). The essential feature of this model is that the signaling components (e.g. Src-family of protein tyrosine phosphatases [SFKs], shown as a yellow circle) move with respect to the linking proteins, which are moving rearward due to actin rearward movement (represented by theyellow arrow in B and C). B. On rigid ECM substrates, the ECM resists unfolding (shown as a thick blue line on the ECM molecule) due the forces of actin rearward movement. Consequently, a quick rise in force near the cell edge is placed onto the linking proteins (shown by the red coil), which causes unfolding and exposure of sites that can be modified (e.g phosphorylated) by the signaling module components near the adhesion. Modifications to the linking module supports additional protein binding and strengthening of adhesions (shown as a light pink oval). C. On soft ECM substrates, the ECM components are unfolded/stretched due to the forces of actin rearward movement. ECM unfolding correlates to a slower rise in force and delayed unfolding of the linking module components until a point further from the cell edge. Because the signal module components remain at the cell edge, they are displaced relative to the adhesion site and are not available to modify the linking module components to promote reinforcement.
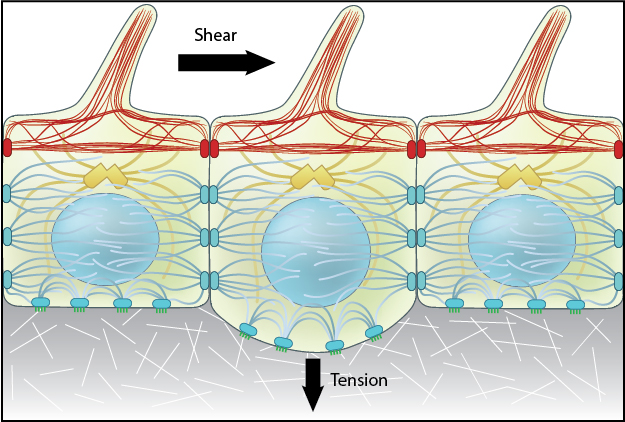
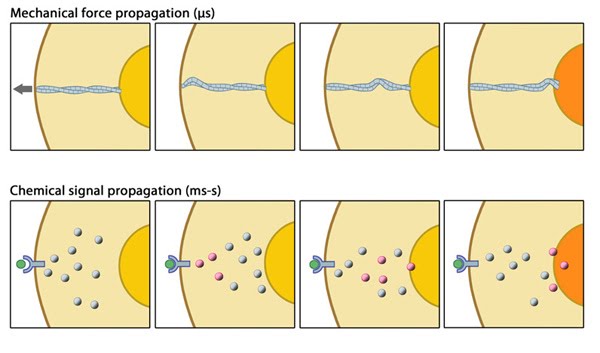
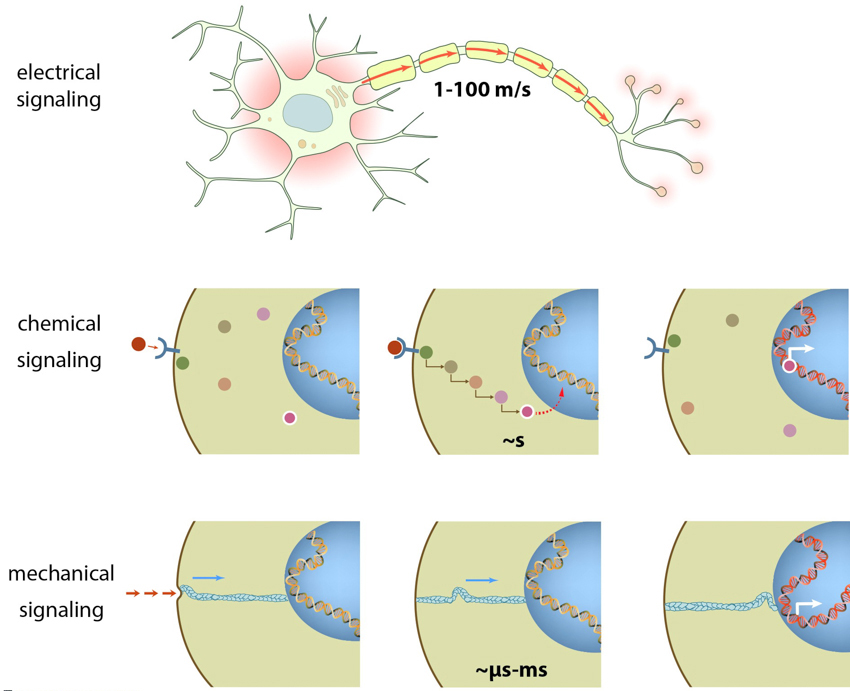
Mechanical versus chemical signal propagation: Mechanical versus chemical signal propagation. Top- Mechanical stress, whether applied to cytoskeleton-linked receptor or generated due to contractility, reaches the nucleus in less than 5 microseconds. Arrow shows direct of tension applied and propagation is depicted as a mechanical wave across the cytoskeletal networks. Bottom-Biochemical signals that originate at the membrane take tens of seconds to travel through their linked network and cause a change in the nucleus (e.g. chromosome remodeling, activate/deactivate gene expression- depicted by color change of nucleus). Adapted from [21508987],[15707995].
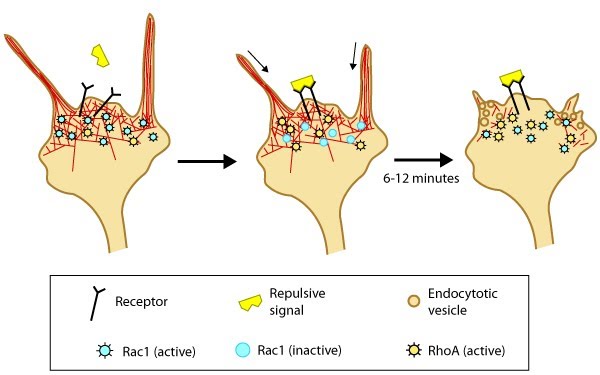
Model of filopodia collapse: In neuronal growth cones, filopodia protrusion stops when a repulsive signal binds to its receptor on the cell surface. Receptor-binding transiently inactivates Rac1 GTPase and prevents it from promoting actin assembly. Resumption of Rac1 activity coincides with filopodia collapse and is required for endocytosis of the collapsing plasma membrane and reorganization of F-actin [32]. RhoA and its effector, ROCK, are activated downstream of repulsive cues [31, 32, 33] and their activity has been implicated in reducing actin polymerization following treatment with repulsive signals [34].
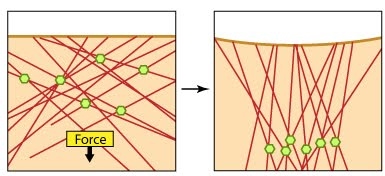
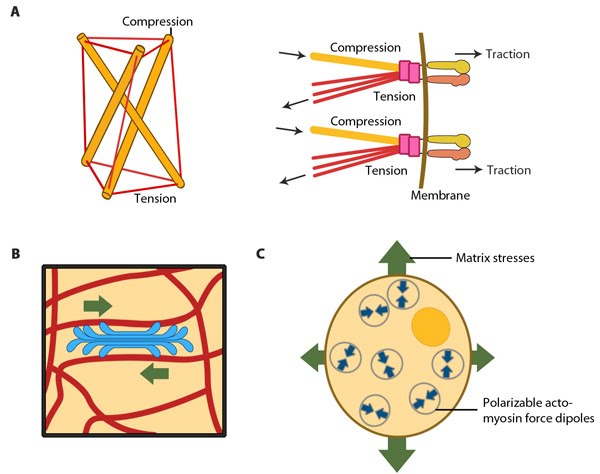
Models for force-induced modulation of cytoskeletal stiffness: Models for force-induced modulation of cytoskeletal stiffness. (A) Tensegrity model: Top left- A simplified version with compression struts and tensed cables exemplifying that stress levels regulate cytoskeletal rigidity. Top right- In the cellular context, microtubules (gold rod) apply compression on cell-matrix adhesions (represented by actin linking modules in pink and integrin dimers) while the actin filaments (red) experience the cellular tension and hence stiffen accordingly. (B) Semiflexible chain model is represented by the flexible actin cables (red) that locally rigidify at points of stress application i.e. myosin (blue bundle) contraction. (C) Dipole polarization model: Formation of contractile actomyosin dipoles is symbolically represented by the arrow pairs. According to this model, they freely orient in response to applied stress as experienced at a particular point. Adapted from Stamenovic D. & Ingber DE. Tensegrity-guided self assembly: from molecules to living cells. Soft Matter 2009; 5:1137–45. [DOI: 10.1039/B903916N] and [20816234].
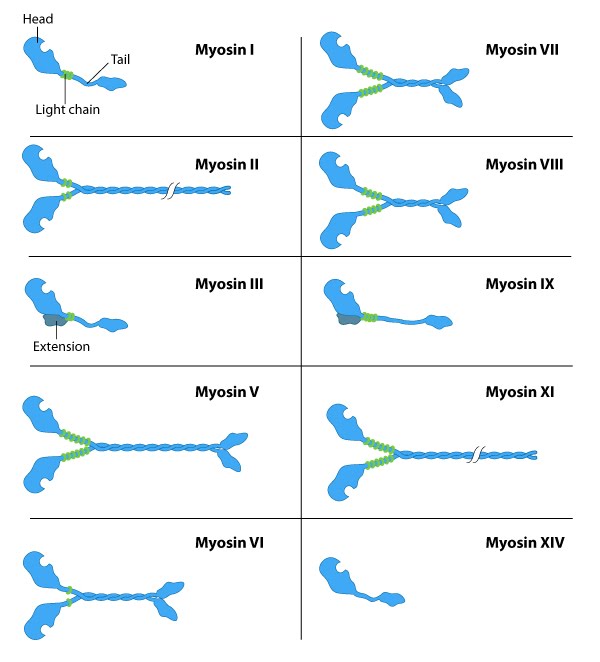
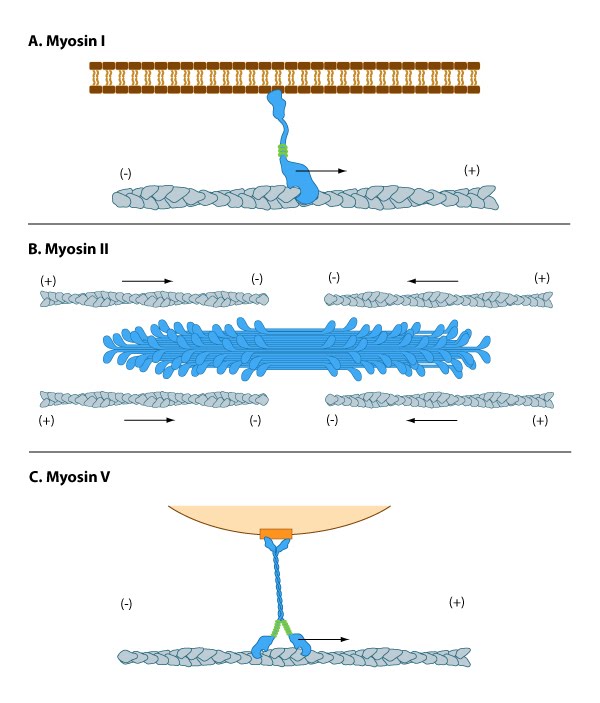
The myosin superfamily of motor proteins: All myosins share a motor domain on their heavy chains at the amino-terminus (the 'head' domain), but they differ considerably at their carboxy-terminus (the 'tail' domain). A few myosin types also have an amino-terminal extension. The number of light chains varies considerably between myosin types and certain myosins exist as dimers. Myosins that form dimers have two motor domains, and the number of light chains can influence the "lever arm" length between the myosin heads – this regulates the length of the myosin 'powerstroke' and the distance the myosin can travel along the actin filament in a single round of ATP hydrolysis (see also 'myosin powerstroke').
111.
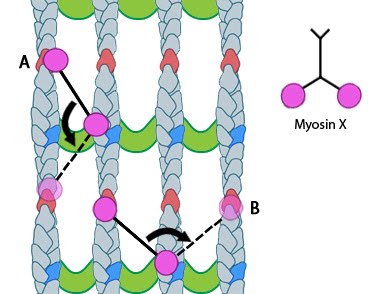
Myosin-X step size: Myosin-X step size corresponds to a single twist of the actin filament helix. Binding sites are represented by red and dark blue actin monomers. In this figure we see how Myosin-X is able to step forwards and backwards (A) as well as horizontally, between filaments (B)
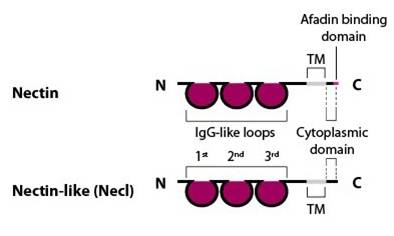
Nectin and nectin-like (Necl) structure: Nectin family members (nectin-1, -2 , -3 and -4) comprise an extracellular domain with three Ig-like loops, a transmembrane domain and a cytoplasmic tail capable of binding the actin-binding protein afadin via a four amino acid conserved motif (Glu/Ala-X-Tyr-Val). Nectin-like family members (Necl-1, -2, -3, -4 and -5) share a similar structure, but are unable to bind afadin [Takai Y, et al, 2003].
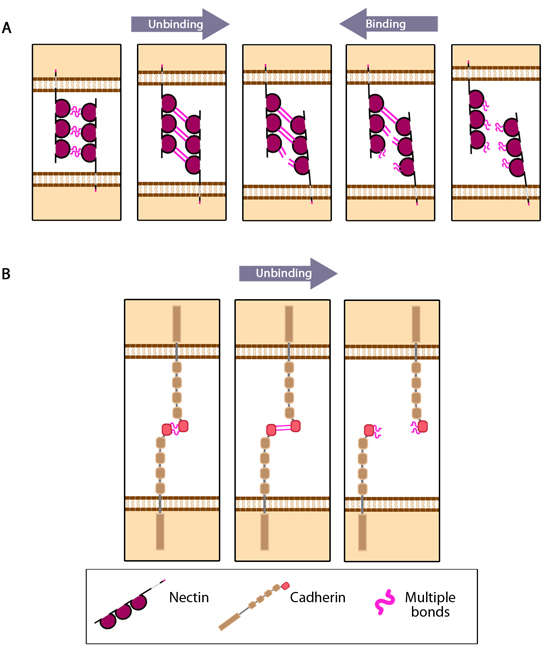
nectin_cadherin_binding_properties: Adapted from [Tsukasaki Y, 2007]. For clarity, single nectin molecules are shown on each cell surface, however it is established that cis interactions of two or more nectin molecules is required before intercellular trans interactions can occur. (A) Multiple bonds nectin-nectin domains are uncooperative in nature resulting in 'zipper-like' binding and unbinding. (B) Multiple bonds between cadherin-cadherin domains are cooperative in nature and longer-lived compared to those bonds between nectin domains on apposing cells. Consequently these interactions unbind in a parallel manner.
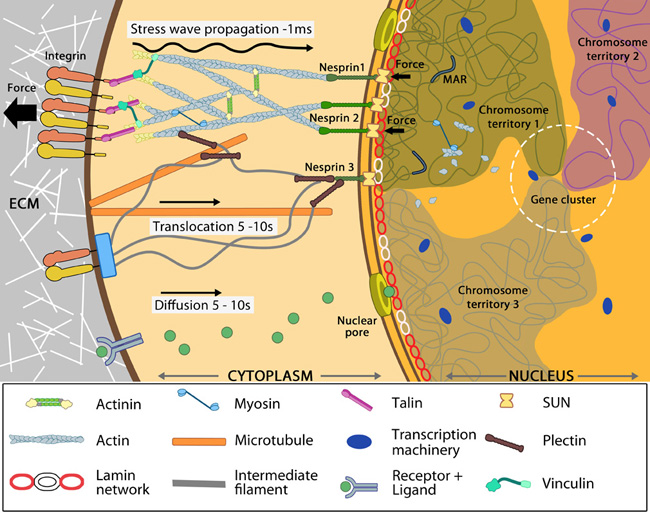
Nuclear connectivity and mechanotransduction: Force experienced by integrins at the cell surface via mechanosensing structures like focal adhesions (integrin cluster linked to actin network), hemidesmosomes (blue rectangle) or cell-cell contact (not shown) is accumulated, channeled through SUN1/SUN2 form the LINC (linker of nucleoskeleton and cytoskeleton) complexes connecting further to the nuclear lamina (red and white lamin network) and hence the attached nuclear scaffold proteins (actin and myosin). Chromatin attaches directly to the lamina and to other scaffolding proteins through the matrix attachment regions (MARs). Upon sensing the force, the nuclear scaffold help repositioning the chromatin thus affecting nuclear prestress and activating genes within milliseconds. Spatial segregation of chromosomes with defined territories is represented as colored compartments inside the nucleus. The dotted circle highlights looping of genes from different chromosomes to form a cluster in 3D space and share transcription apparatus (navy ovals). On the contrary, chemical signaling mediated by motor-based translocation along cytoskeletal filaments or diffusion of activated regulatory factors takes few seconds. Adapted from [19197334, 21391812]
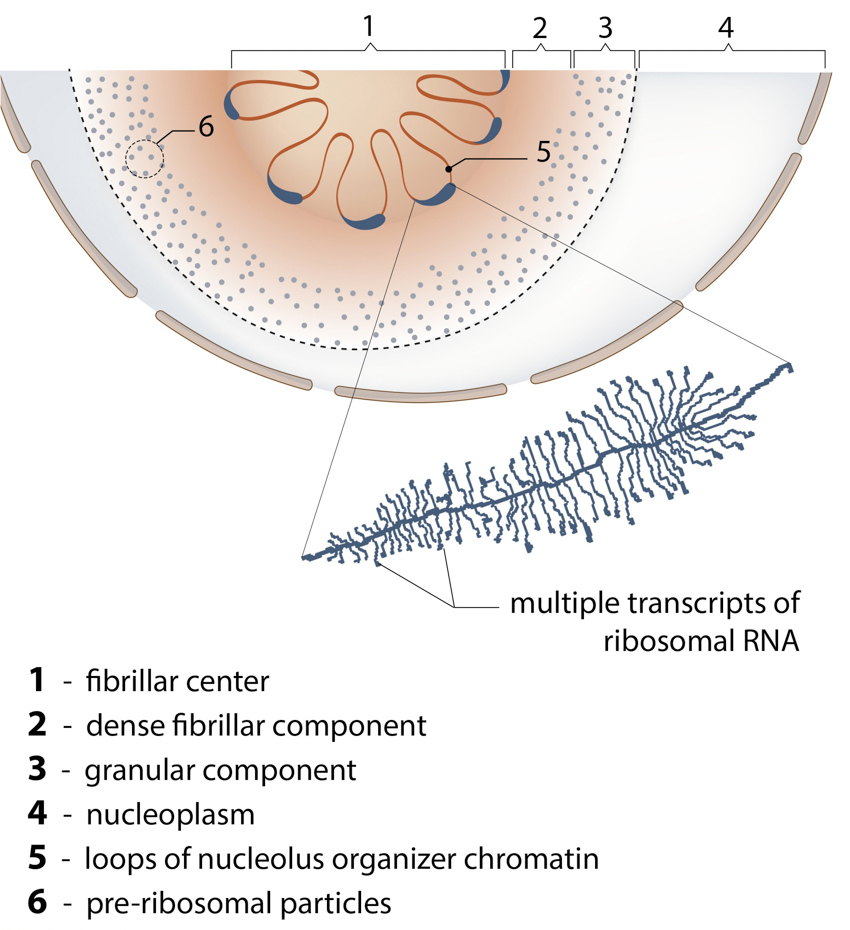
The Nucleolus: The main function of nucleolus is the synthesis of ribosomal RNA and assembly of ribosomal particles. Nucleoli are often enriched with centromeric satellite repeats and inactive gene clusters.
Nucleoskeleton stabilizes the nuclear structure.: The nuclear lamina and associated proteins provide structural integrity to the nucleus. While the lamina confers stiffness in particular, the accessory structural proteins are responsible for other mechanical properties such as resilience. Emerin connects lamins, LINC complexes and the other nucleoskeletal components at the nuclear interior. Spectrin is involved in organizing the lamins and hence provide overall integrity. This is also aided by protein 4.1 and small actin filaments that act as mechanical struts to support the links. Adapted from [21321324, 15688064].
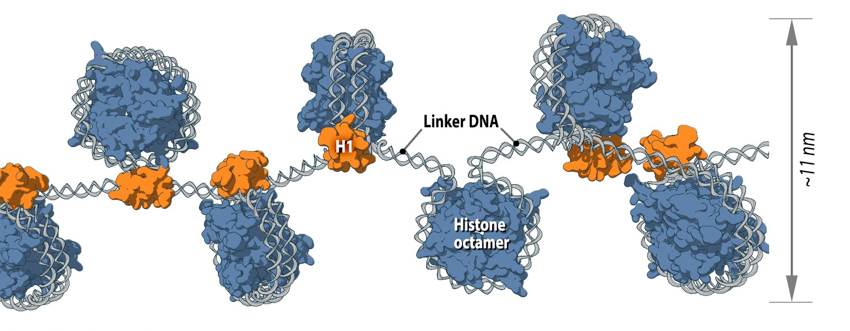
Nucleosome is the first level of DNA packaging:: Each nucleosome consists of histone octamer core, assembled from the histones H2A, H2B, H3 and H4 (or other histone variants in some cases) and a segment of DNA that wraps around the histone core. Adjacent nucleosomes are connected via “linker DNA”.
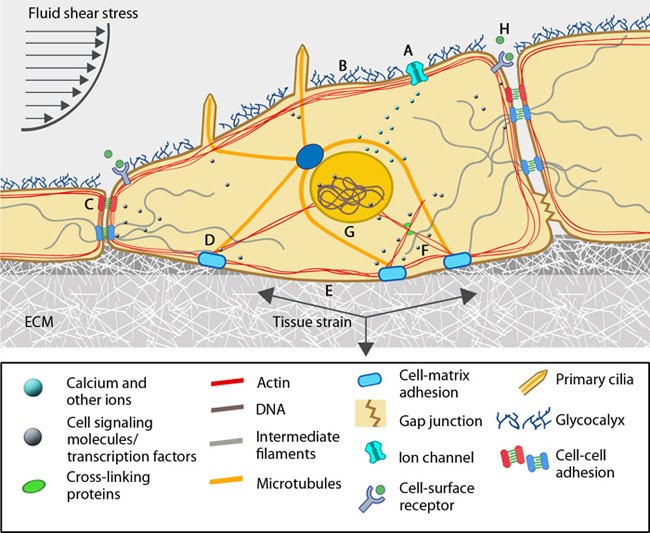
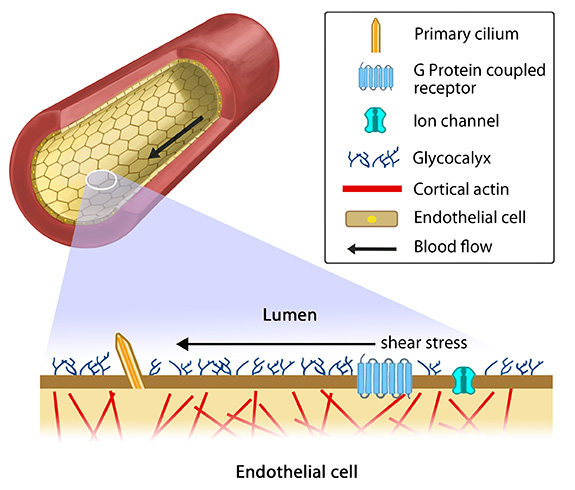
Overview of mechanotransduction in a cell: Adapted from [19197333]. Mechanotransduction and response are often mediated by several overlapping signaling pathways. A variety of cellular components that are suggested to acts as mechanosensors and force transducers are depicted in a representative cell. Some of the features are specific to cell types. (A) Ion channels get activated in response to lipid fluidity and stretching of the plasma membrane to allow ion flow. Additionally changes in G-protein coupled receptors can induce intracellular signals. (B) A layer of glycoproteins found on the surface of endothelial cells, namely glycocalyx, have the ability to sense fluid shear. (C) Cell-cell adhesion complexes: Adherens junctions (red) and desmosomes (pale blue) serve as cytoskeletal communicative conduits while gap junctions connect the cytoplasm of adjacent cells. (D) Cell-matrix adhesion complexes probe the cellular environment, sense and signal the changes in the extracellular mileu. (E) Force-induced conformational changes in the ECM components can sensitize the receptors and initiate mechanotransduction signals. (F) Intracellular strain changes protein conformations and hence the binding capacity of cytoskeletal components, their crosslinkers and motor proteins to other molecules leading to alterations in signaling pathways. (G) The nucleus is proposed to sense the mechanical signals and accordingly modulate the transcription machinery by changing chromosome positions, chromatin arrangements and transport of molecules across the nuclear membrane. (H) It is to be noted that changes in inter-cellular space is cell-type specific. This can influence concentration of signaling molecules/ proteins that bind cell-surface receptors.
121.
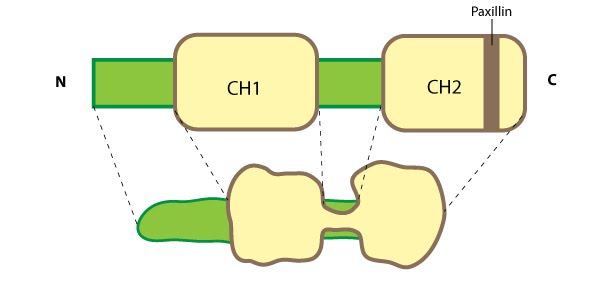
Parvin: This schematic diagram illustrates the molecular organization of parvin and provides examples for how parvin is presented in figures throughout this resource. Relevant domains believed to be important for binding to actin (e.g. CH1, CH2) and protein-protein interactions are highlighted (reviewed in [16314921])
122.
Paxillin: This schematic diagram illustrates the molecular organization of paxillin and provides examples for how paxillin is represented in figures throughout this resource. Relevant domains believed to be important for protein-protein interactions are highlighted (reviewed in [7]). These interactions include FAK, vinculin [8, 9], Src [10], parvin [11],tubulin [12], α-integrin [13, 14, 15], and focal adhesion targeting [16].
123.
Paxillin Localization: A HFF (human foreskin fibroblast) cell plated on a fibronectin coated glass coverslip, transfected with GFP-LifeAct (green), which labels F-actin in living cells, and mCherry-paxillin (red). It was imaged on Olympus IX81 Inverted microscope using a Perkin Elmer spinning disk at 100x magnification. Image courtesy: Yee Han Tee, Mechanobiology Institute, Singapore.
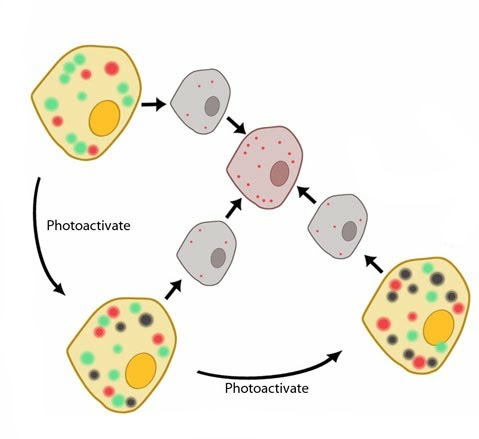
Photoactivated Localization Microscopy (PALM): PALM is a method of super-resolution microscopy where repeated rounds of excitation, emission and photobleaching are required to produce a single super-resolution image. Each round affects only a fraction of the total flurophores present in the sample. This means that fluorophores in close proximity to each other may be individually resolved. This is depicted in this diagram where unaffected fluorophores appear green, some flurophores are excited and emit fluorescence (red), whilst others are photobleached from previous rounds of excitation (black). Imaging only the few-optically resolvable fluorophores (red) in each round means the data can later be combined to reconstruct a super-resolution microscopy image.
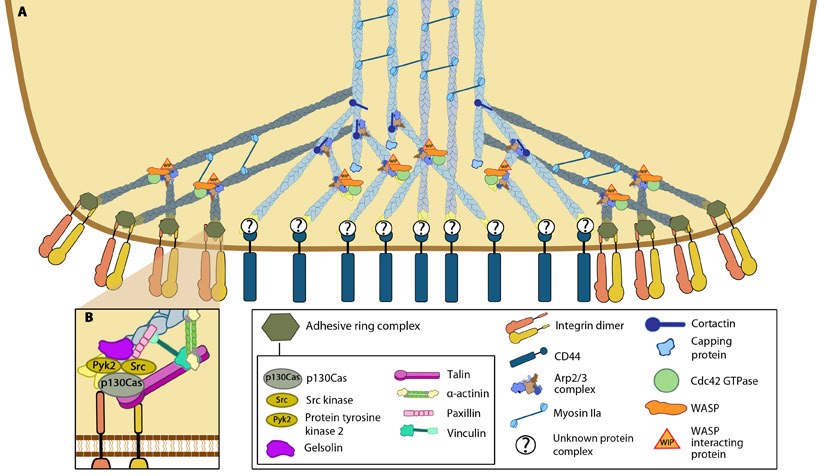
Podosome structure: (A) The actin core of the podosome (light blue) includes several proteins that promote actin nucleation. The Arp2/3-Cdc42-WASP-WIP complex facilitates actin nucleation and branching proximal to the plasma membrane, whilst cortactin (or its leukocyte-specific homologue HS-1[21398607]) acts to stabilize branch points more distally [21785274]. It is not yet clear whether the attachment of the actin core to CD44 receptors occurs via branched or unbranched actin filaments or what protein complex could facilitate this link. Radial actin filaments (dark blue) emanate from the actin core [ 21779330] and link to cell surface integrins that form the basis of the adhesive ring complex. The radial network is speculated to exhibit mechanosensory potential [18760605], involving the transmission of forces in a myosin II dependent manner. (B) The ring complex is formed by a circular array of integrins, to which several other proteins also localize. These proteins include the signaling proteins, Src kinase and Pyk2, the adaptor protein p130CAS, the actin severing protein gelsolin and the focal adhesion-associated proteins talin, vinculin, paxillin and α-actinin. It should be noted that though gelsolin localizes to this ring structure, it is not absolutely required for podosome formation [21779330].
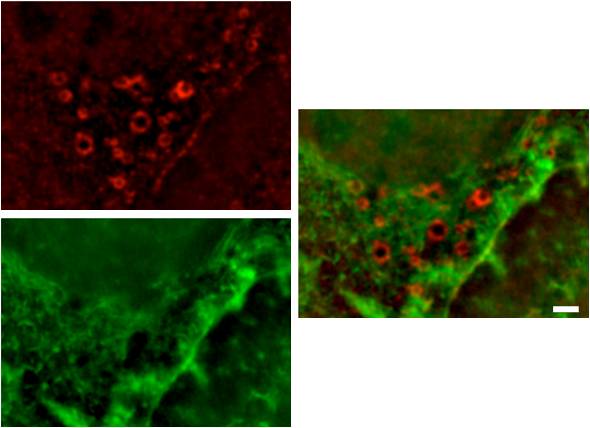
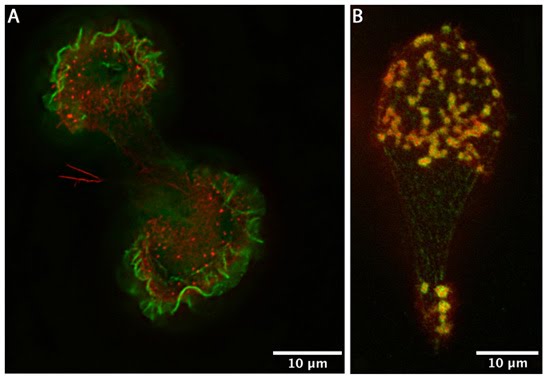
Podosomes: (A) Human acute monocytic leukemia cells (THP1) cultured on fibronectin, differentiated towards macrophages with TGF beta1, displaying podosomes (red puncta). Actin was visualized using phalloidin-TRITC. The image is a projection of several Z-planes where lamellipodial actin ruffles on upper planes are false-colored green and actin cores of podosomes on lower planes are inred. (B) Human acute monocytic leukemia cells (THP1) cultured on fibronectin, differentiated towards macrophages with TGF beta1, displaying podosomes (yellow puncta). Actin (red) was stained usingphalloidin-TRITC and vinculin (green) using Alexafluor488-coupled anti-vinculin antibody. The actin core and the surrounding ring complex (vinculin) are clearly visible. The cells were imaged using a DeltaVision microscope. [Images captured by Tee Yee Han, Mechanobiology Institute, Singapore]
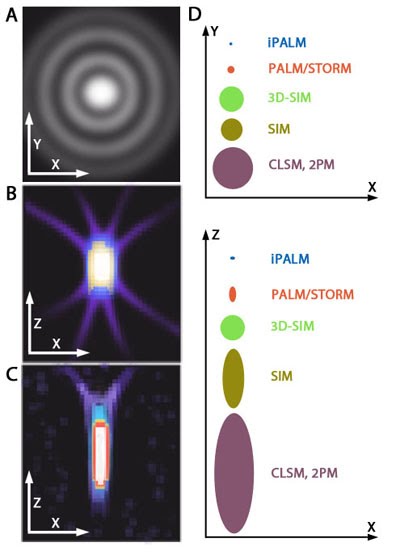
PSF elongation along the Z axis.: Image adapted from [6] When imaging along the Z axis (e.g. the depth of the cell body), PSF elongation is one factor that lowers the effective resolution of standard light/fluorescence microscopy. (A) A representation of an ideal diffraction pattern in the X-Y plane. (B)and (C) Representative images highlighting symmetrical side projections in the X and Z axis, as well as elongated distribution and asymmetry along the Z-axis. These images are based on microscopy images of fluorescent beads in aqueous solution (A) and a collagen gel ((B) and (C)). (D) Graphs depicting the extent of PSF elongation along the z axis when compared to the x-y plane using different types of microscopy. Elongation is most evident in confocal laser scanning microscopy (CLSM). 3D-SIM overcomes PSF elongation along the Z-axis compared to traditional SIM, whilst iPALM greatly refines the PSF, to the point that it is slightly smaller along the Z-axis compared to the X-axis.
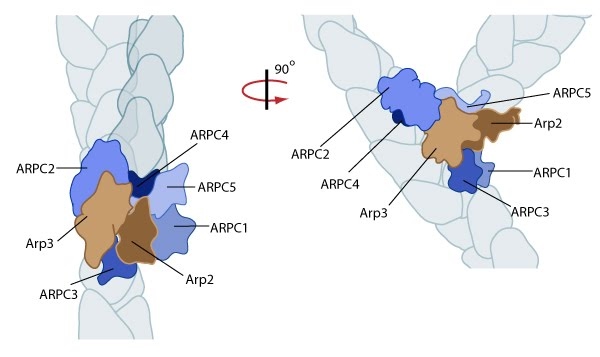
Reconstruction of Arp2/3 complex-mediated nucleation: Arp2/3 complex initiates a new branched filament by binding to the side of the mother filament and recruiting actin monomers. Components of the Arp2/3 complex remain at the pointed end of the filament (Figure adapted from [18316411]).
133.
Regulation of actin treadmilling: The length of actin filaments is controlled by actin binding proteins. Capping proteins prevent assembly at the barbed end while ADF binds to the side of ADP-actin filaments to cause disassembly of the filament. In the absence of actin-binding proteins, the filament length is stable by a treadmilling mechanism (middle filament). Profilin enhances filament assembly by promoting ADP to ATP exchange on actin and by directing actin monomers to the barbed end of filaments (bottom filament). (Figure adapted from [8]).
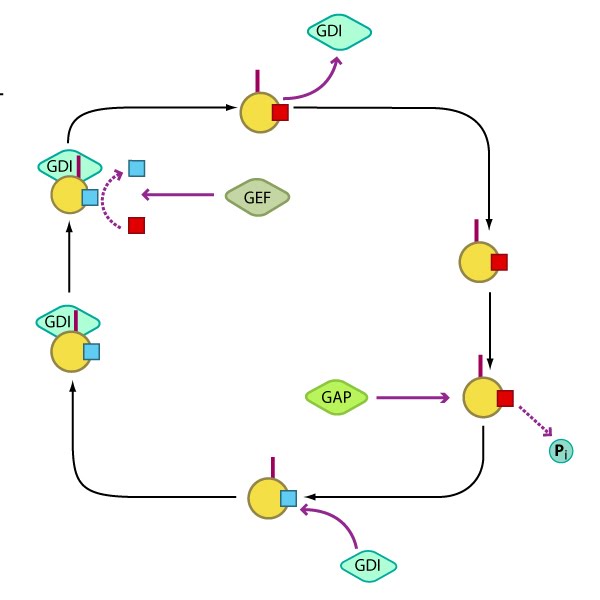
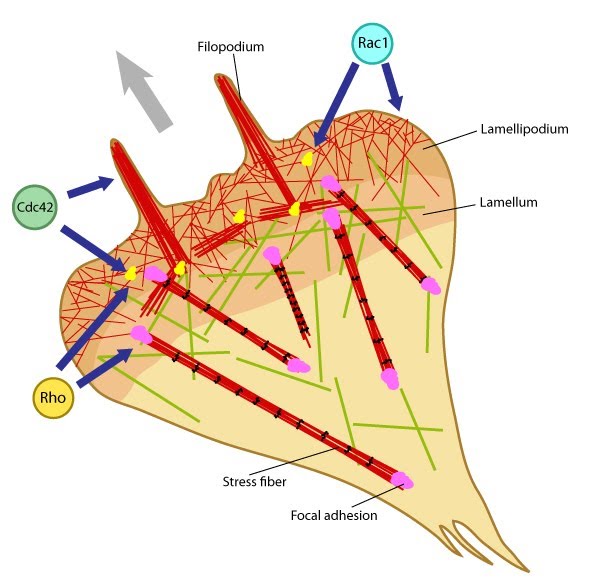
Rho GTPases regulate adhesion dynamics and cytoskeletal reorganization during motility: Rho family GTPases Cdc42, Rac1 and Rho act at different regions in a cell (indicated by arrows) to orchestrate migration. Cdc42 generally controls the cell polarity and the formation of filopodia and nascent focal adhesions (shown as yellow dots). Rho influences cell adhesion assembly and maturation, in addition to controlling stress fiber formation and contractile activity. Rac1 primarily controls actin assembly and nascent adhesion formation in the lamellipodium.
136.
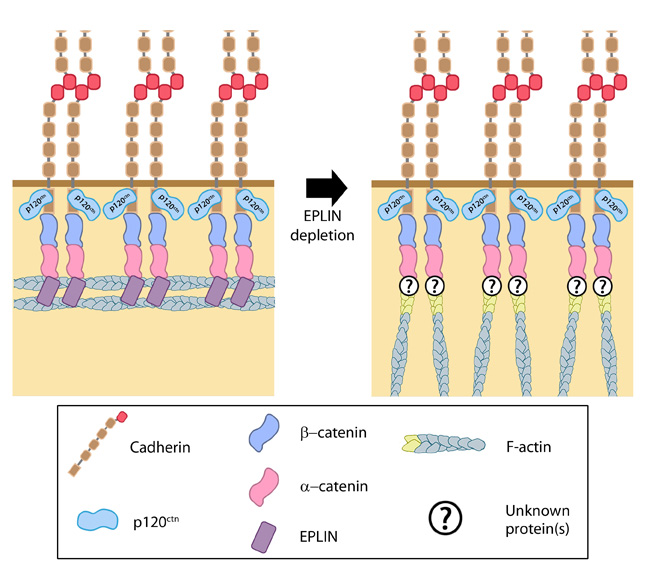
Role of EPLIN in adherens junctions: Adapted from [33].EPLIN is able to bind α-catenin, bound to the β-catenin-cadherin complex, as well as actin. It is therefore suggested as a candidate for linking the cadherin-catenin complex to the actin cytoskeleton. Moreover, it is also suggested to promote and stablize formation of the adhesion belt, with its loss causing the actin cytoskeleton in this region to take on a more immature appearance of radial filaments [31].
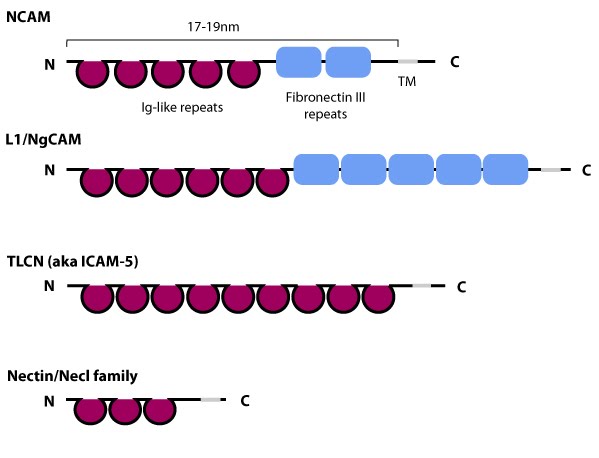
Schematic diagram of immunoglobulin superfamily members that are found in neurons: The extracytoplasmic portion of all Ig superfamily members is composed of a variable number of Ig domains (~3.7 nm per Ig domain) that together yield a final length of approximately 17-19 nm for both NCAM [3] and ICAM-1 [2]; certain Ig family members also contain a variable number of fibronectin III repeat domains. The transmembrane domain (TM) located at the extreme carboxy-terminus leaves a relatively small portion on the intracellular side for associating with other proteins. ICAM-1 is similar to ICAM-5 (not shown) but contains five Ig domains. Nectin and Necls share a similar structure, with three extracytoplasmic Ig-like domains (reviewed in [4])
Signal transduction:: Cells sense their environment through distinct mechanisms and subcellular structures. These allow cells to produce and detect a variety of signal types, from soluble molecules or chemicals, to electrical current and even mechanical cues that are transmitted via the push and pull of biomolecules.
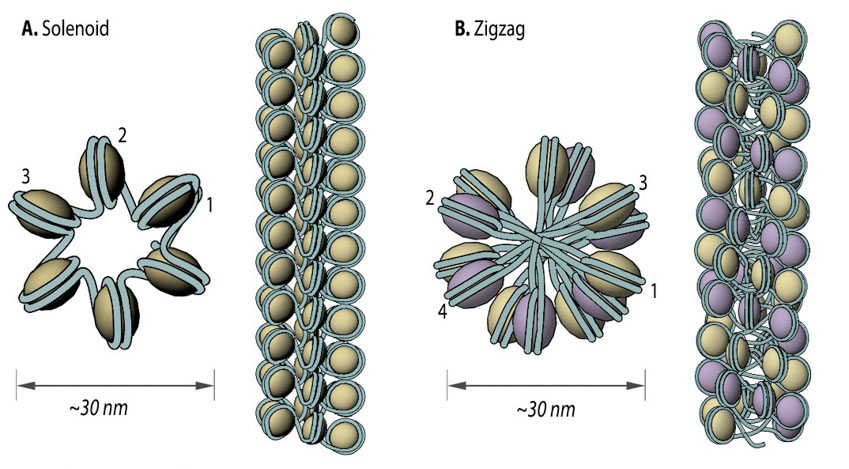
Solenoid (A) and zigzag (B) models of intermediate chromatin condensation.: 30 nm chromatin fibers are considered to exist in the form of so called solenoid or zigzag. The main feature of solenoid model is that nucleosomes follow each other along the same helical path, and interactions between the histone cores occur sequentially (1, 2, 3 and so on). Therefore, solenoid is also referred to as “one start model”. In zigzag, on the other hand, linker DNA connects two opposing nucleosomes, creating a structure where the alternate histone cores become interacting partners (i.e., 1 and 3, 2 and 4 and so on). Therefore, zigzag is considered as a “two start model”, which is indicated in the figure (B) by two different colors of histone cores: yellow interacting nucleosome partners (1, 3, etc.) as opposed to the violet nucleosome row (2, 4, etc.).
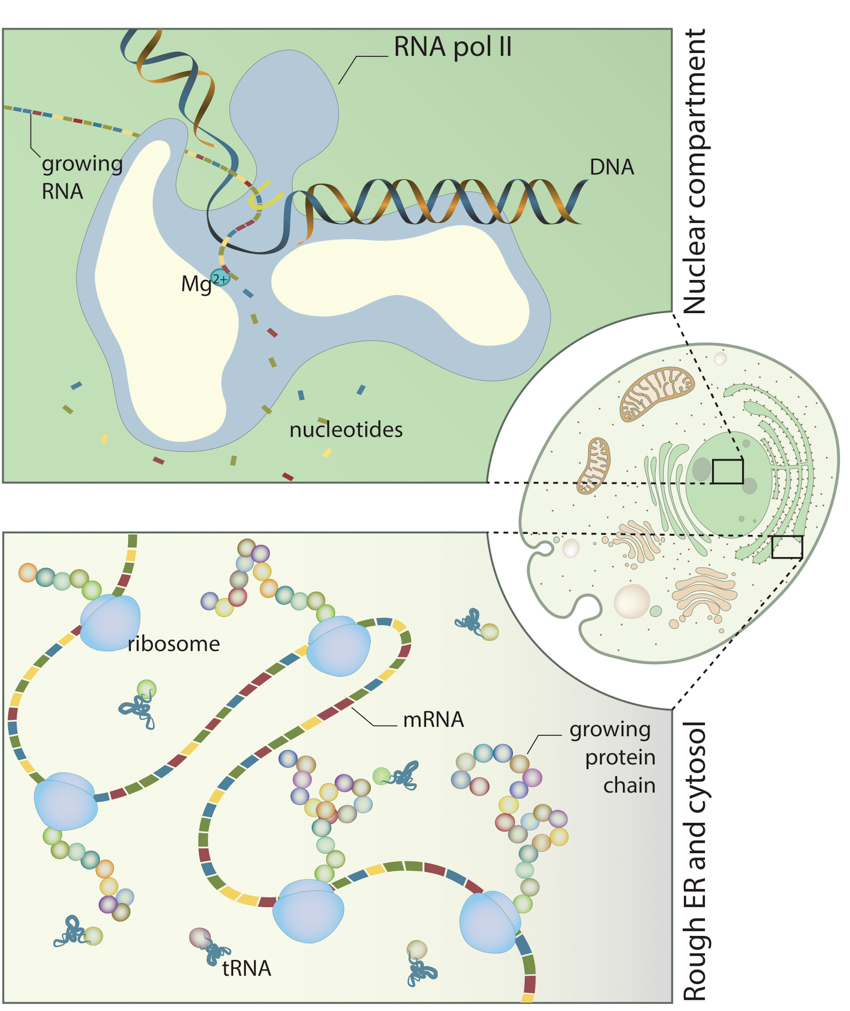
Spatiotemporal control of synthesis:: DNA and RNA are synthesized in the nucleus (upper panel). RNA is then transferred into the cytosol or rough endoplasmic reticulum (lower panel), where protein synthesis takes place.
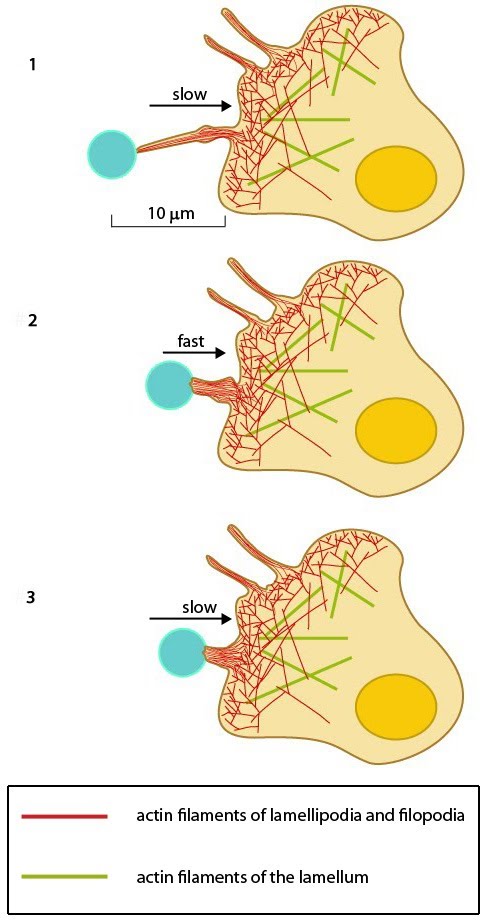
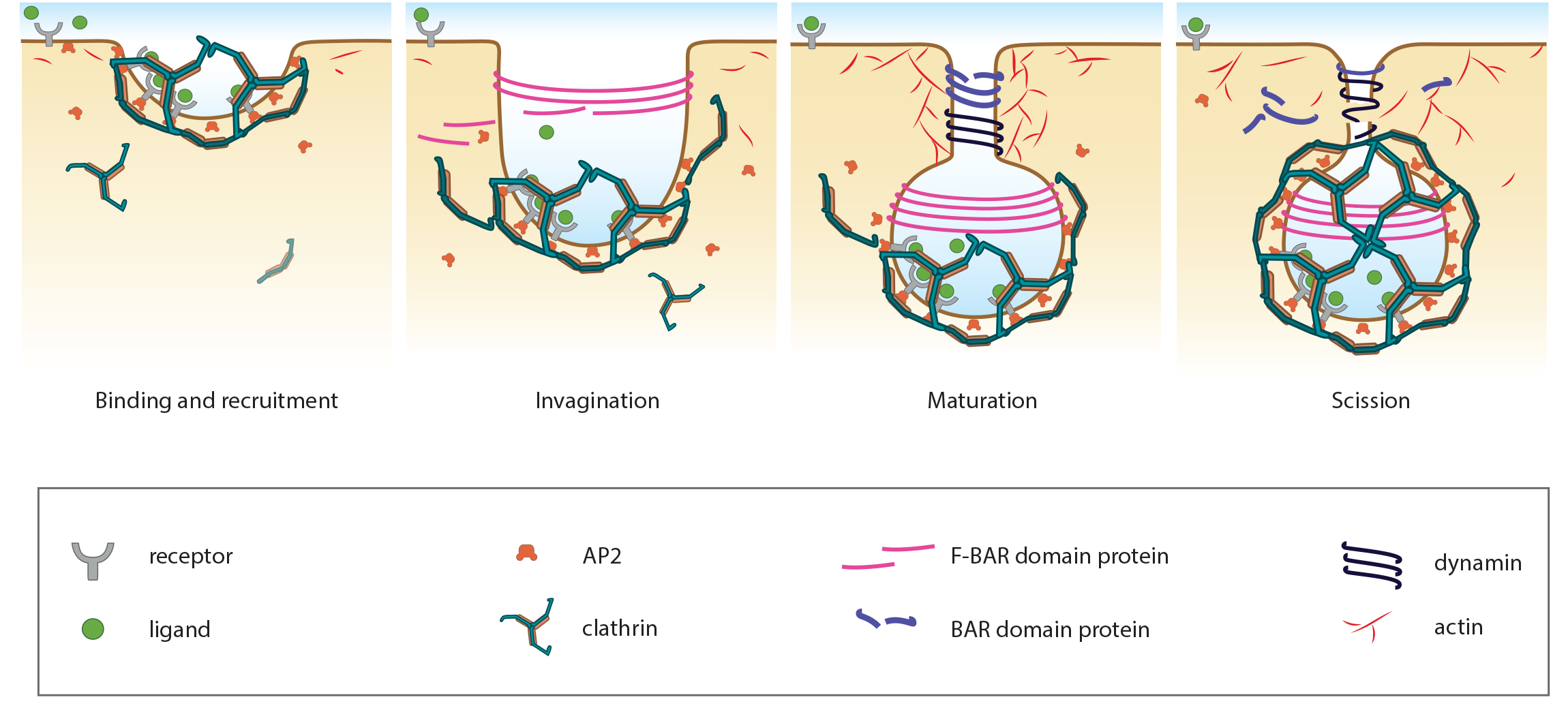
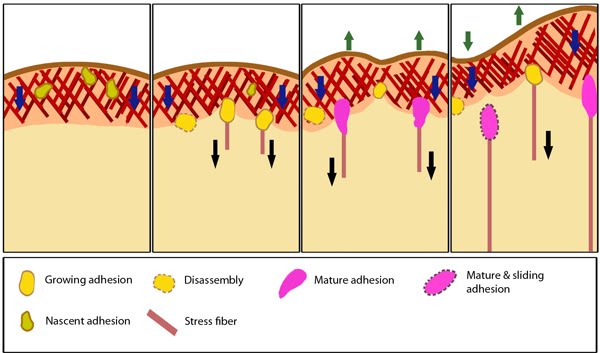
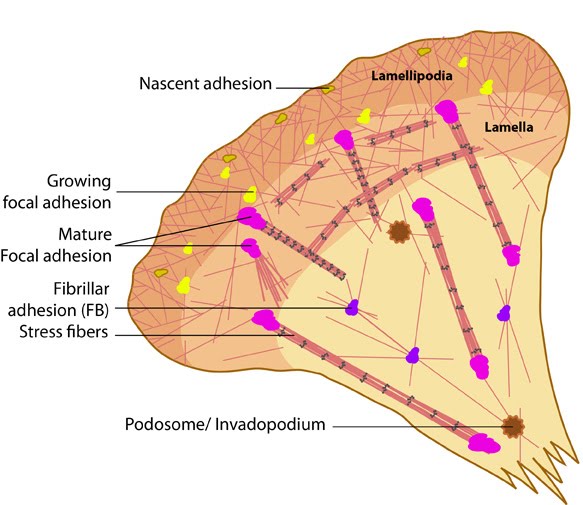
Stages in focal adhesion formation: The four panels represent roughly the different steps in the formation and disassembly of focal adhesions. The lamellipodium is distinguished from the lamellum by a color difference; the actin filaments represent the retrograde flow and the blue arrows indicate its characteristic centripetal flow. From left: Panel 1 shows formation of nascent adhesions at the cell periphery. In panel 2, some adhesions attach to stress fibers and grow in size while some disassemble (yellow with dotted outline) at the lamellipodium-lamellum boundary. Maturation happens upon increase in tension along the stress fibers (panel 3). The protrusions of the cell edge are shown as green arrows corresponding to points of adhesion maturation (panels 3 and 4). Panel 4 shows sliding of adhesion and hence slight retraction of the respective section of the cell edge compared to panel 3. Decrease in retrograde flow at points of adhesions is marked by the dipping of the lamellipodial border. Adapted from [4, 8].
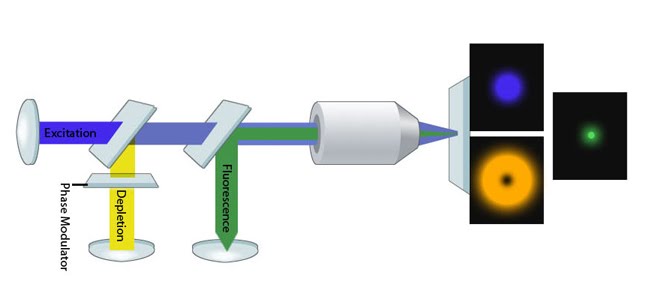
STED Microscopy Schematic: Image adapted from [2]. An excitation beam (blue) is surrounded by a depletion (de-excitation/STED) beam (yellow). Fluorophores that lie in the path of the depletion beam (those in the 'donut') return to the ground state immediately whilst those molecules within the center of the PSF are exposed only to the excitation beam and hence produce a fluorescent signal (green).
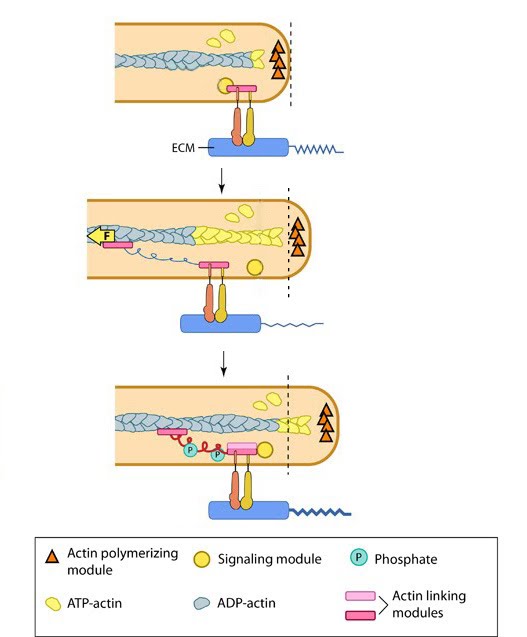
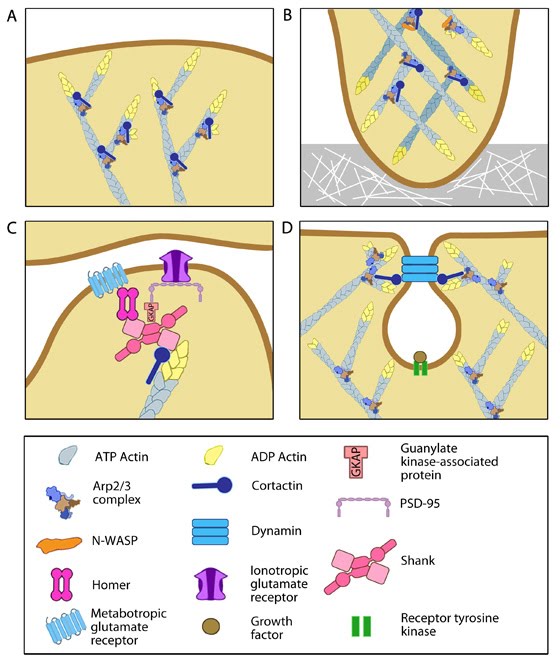
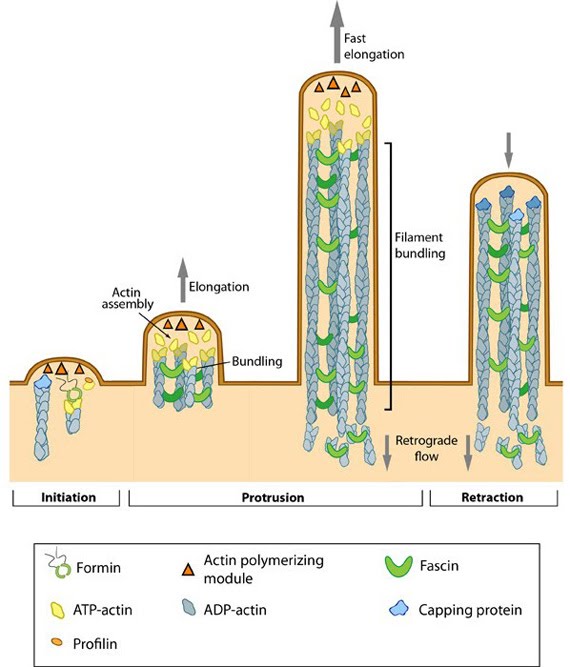
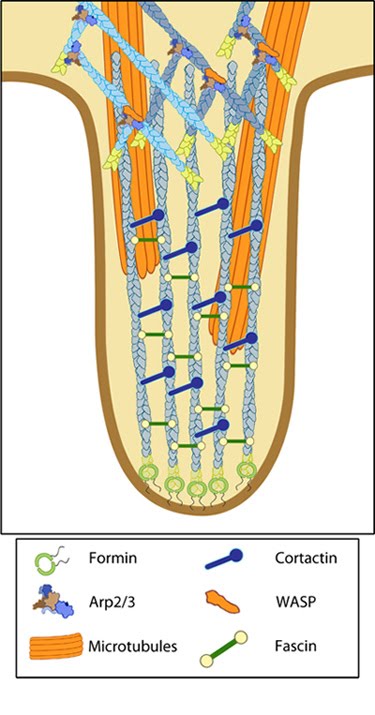
Steps in filopodium formation: Steps in filopodium formation. Actin filament assembly can be initiated by uncapping pre-formed actin filaments or by de novo formation (which includes both formin- and Arp2/3-mediated [not shown] nucleation). The force produced by actin assembly at the barbed end of actin filaments drives membrane protrusion. Numerous proteins (including IRSp53, Ena/VASP proteins, WASp/Scar proteins) cooperate to promote actin-assembly and enhance bundling of actin filaments by fascin. When the barbed end of the filament is capped, this stops filament assembly and protrusion; continued retrograde movement of the filaments results in retraction of the filopodium. (Figure adapted from Faix J and Rottner K, 2006)
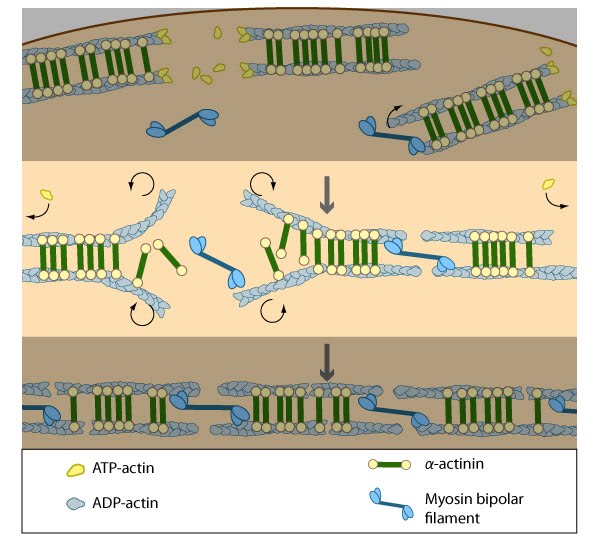
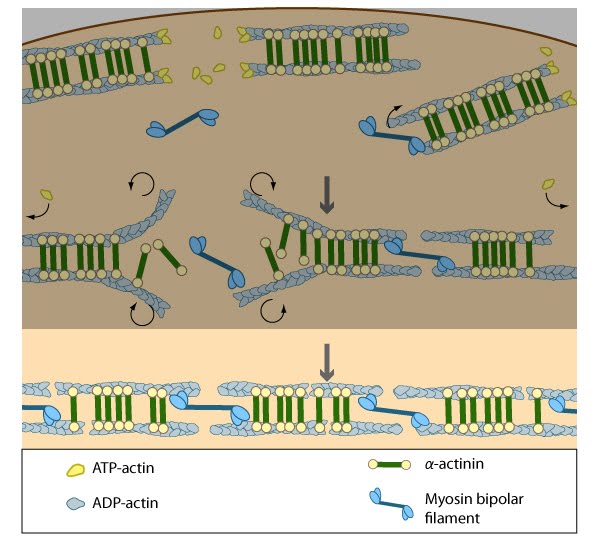
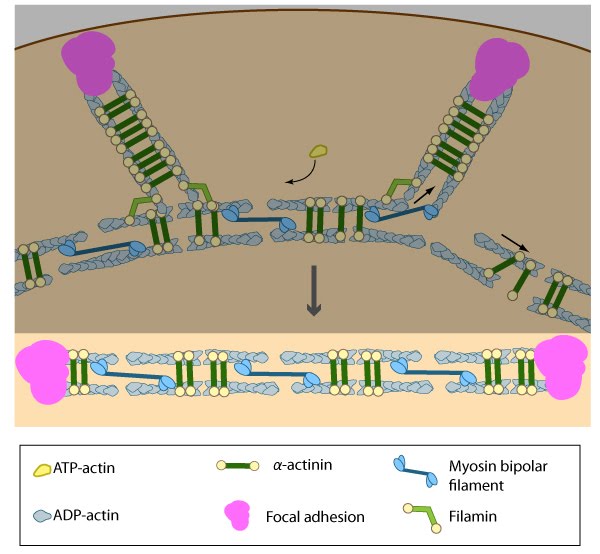
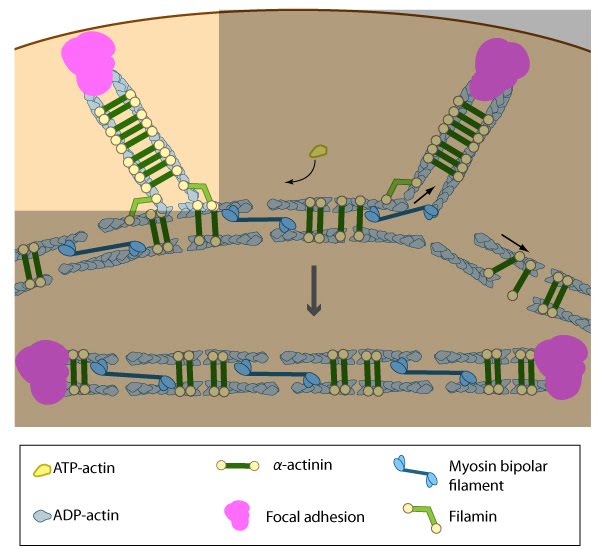
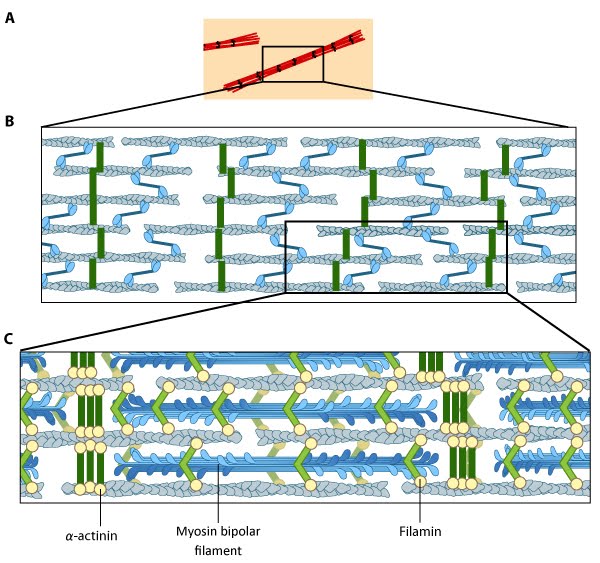
Stress fiber structure: A. Isolated stress fibers have a banded appearance, with bundles of actin filaments interspersed with semiperiodic electron-dense regions. B. The electron-dense regions are rich in actin crosslinking proteins, namely α-actinin. Bipolar myosin II filaments lie between the loosely packed actin filaments in the regions that lack α-actinin . C. A high resolution view of the bipolar myosin filament heads interspersed between the regions rich in α-actinin. Relative to α-actinin, the more flexible actin crosslinking protein, filamin, is dispersed throughout the entire stress fiber.
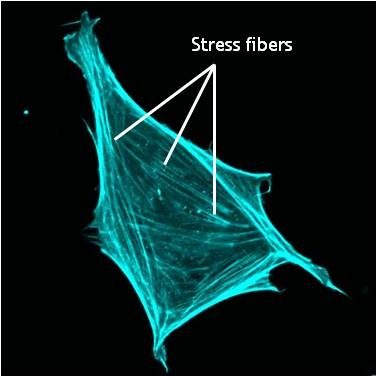
Stress fibers in a cell labeled for F-actin: A mouse embryonic fibroblast of the RPTPa cell line, plated on a fibronectin coated glass cover slip. The cell was transfected with RFP-Lifeact (a kind gift from Dr Roland Wedlich-Soldner, Max Planck Institute of Biochemistry, Germany), which labels F-actin in living cells. The cell was imaged using a Nikon A1Rsi confocal microscope at 60x magnification and false coloured cyan. Image captured by Wei Wei Luo, Mechanobiology Institute, Singapore.
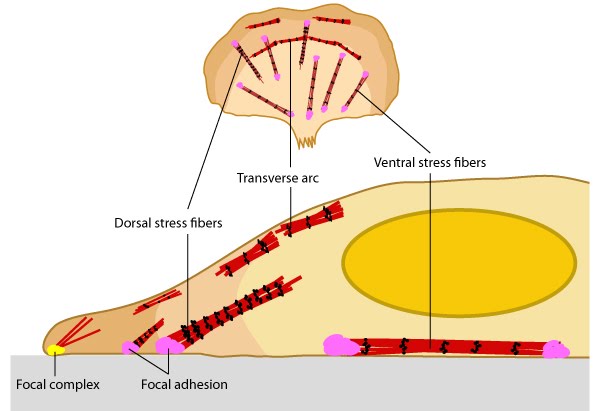
Stress fibers link the cell interior to the exterior through focal adhesions: A. Stress fibers in slowly migrating cells, e.g., bone cancer cells (left) and fibroblasts (right). B. The presence of myosin thick filaments in stress fibers and transverse arcs makes them contractile. C. Dorsal/ventral stress fibers are anchored to the external surface through the focal adhesions. Assembly of over 165 component molecules into the FA is modulated by internal forces from the contraction of myosin and actin filaments in the stress fiber, or external forces generated on ECM components.
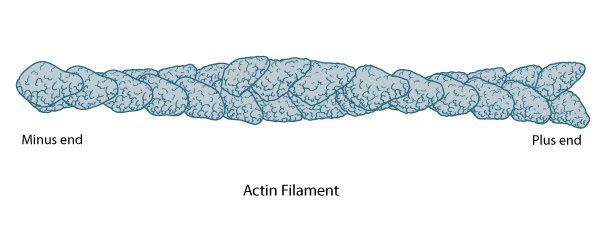
Structure of an actin filament showing the barbed (or plus) and pointed (or minus) ends: This diagram illustrates the molecular organization of actin and provides examples for how an actin filament is represented in figures throughout this resource. Early models for actin filaments were constructed by fitting the filament x-ray crystal structure to the atomic structure of actin monomers [2395461](reviewed in [3897278]) while more recent models use a number of different approaches [17278381].
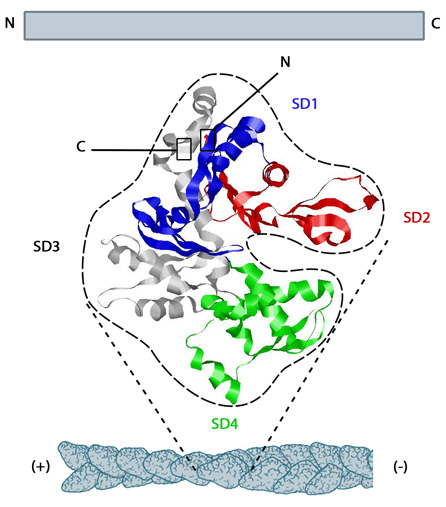
Structure of G-actin and its assembly into filaments: The structure shown here [3] was downloaded from the RSCB Protein Data Bank (PDB file: 1atn). The ATP binding cleft is starred (*) on the right. Actin comprises four subdomains, termed SD1 (blue), SD2 (red), SD3 (grey) and SD4 (green). The barbed end of each monomer (SD1 and SD3) is shown on the left and the pointed end (SD2 and SD4) is shown on the right. Similarly the polarity of the actin filament, which comprises these monomers is shown at the bottom, with the barbed (+) end on the left and the pointed (-) end on the right.
151.
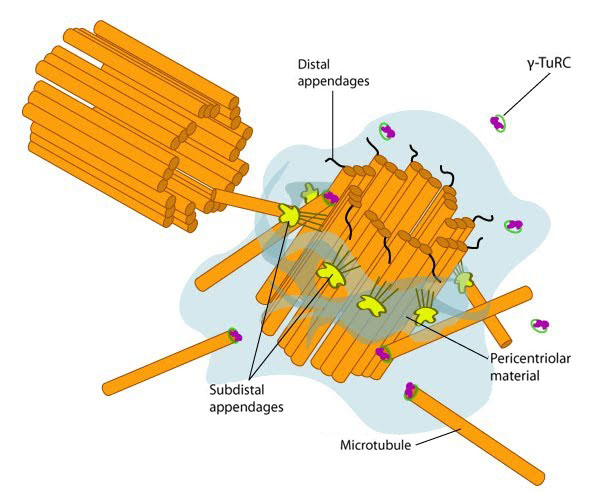
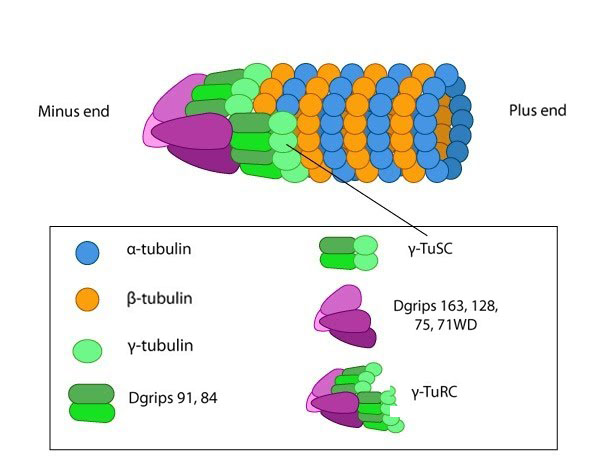
Structure of the centrosome: In non-dividing cells, the centrosome, which is also known as the MTOC, consists of a pair of L-shaped centrioles and associated pericentriolar material. The 'older' of the two centrioles has additional proteins that form appendages along the exterior surface. The pericentriolar material contains numerous γ-TuRCs that nucleate the microtubule array. The centrioles have microtubules organized in a structure similar to the basal bodies found at the base of cilia and flagella. Figure adapted from [10854327]
152.
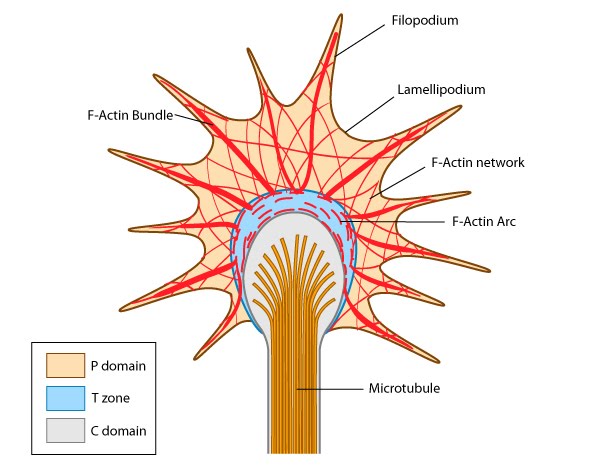
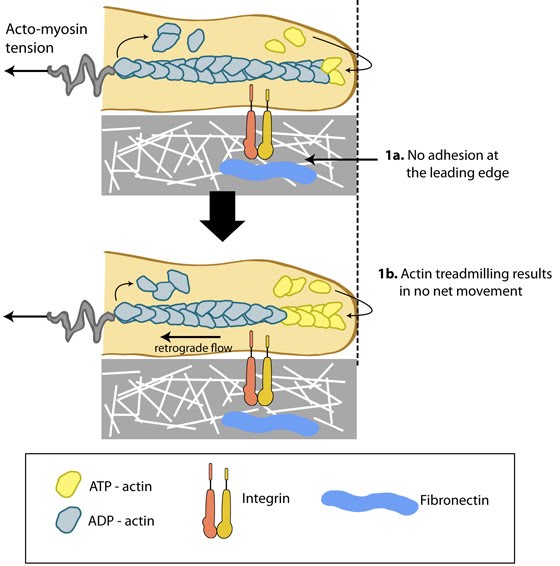
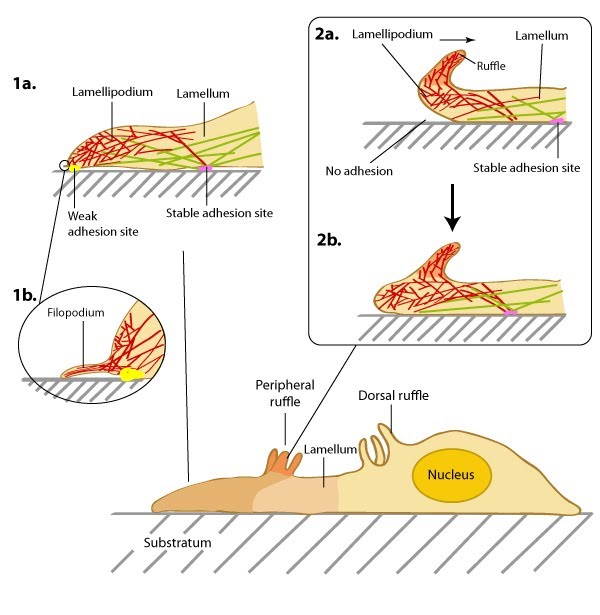
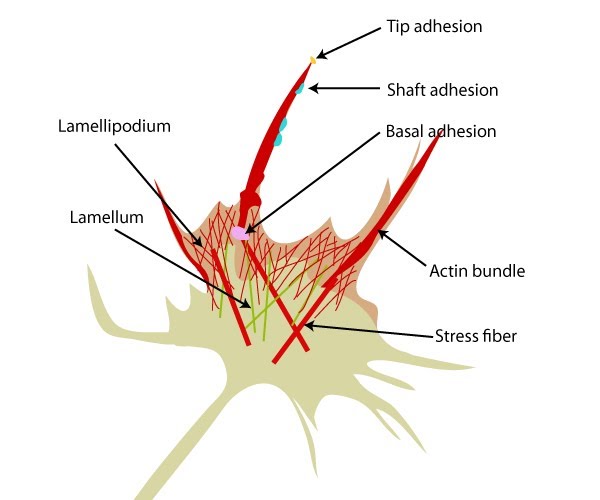
Structure of the lamellipodium and the lamellum: Actin filament assembly in the lamellipodium and retrograde movement of the filaments generates protrusive forces that drive cell motility in a specific direction. Lamellipodium (1a) and filopodium (1b) are common actin-based structures that are used to probe the cellular environment ahead of migrating cells. Adhesion to an underlying surface (ECM, other cells etc) determines the rate and direction of actin assembly during cell spreading and movement. During cell migration, nascent adhesions at the cell front 'mature' into stable adhesions as they progress towards the lamella. In the absence of adhesion at the leading edge (2a, 2b), the retrograde movement of actin filaments is converted into ruffles.
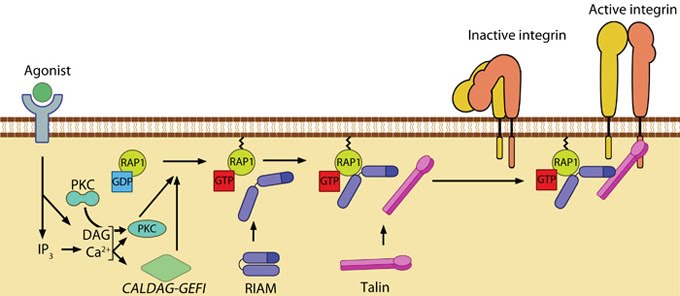
Talin recruitment to membrane: Talin recruitment to membrane. Ligand occupancy in certain cell-surface receptors (agonists) causes phospholipid hydrolysis releasing diacylglycerol (DAG) and inositol triphosphate (IP3). IP3 increases cytosolic levels of calcium ions; DAG and Ca2+ can promote GTP-loading and membrane translocation of Rap1 either by activating Ca2+ and DAG-regulated GEF (CALDAG-GEF or Rap-GEF) or protein kinase C (PKC). Activated Rap1 in turn, recruits Rap1-GTP-interacting adaptor molecule (RIAM) along with its binding partner, talin to the plasma membrane. Adapted from [18573917, 20308986].
154.
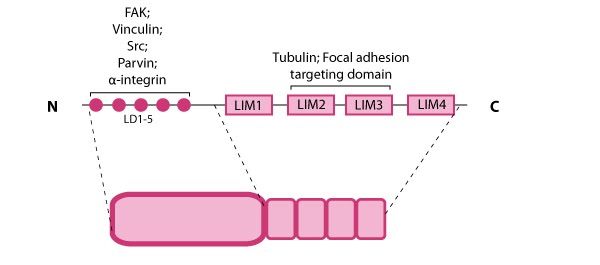
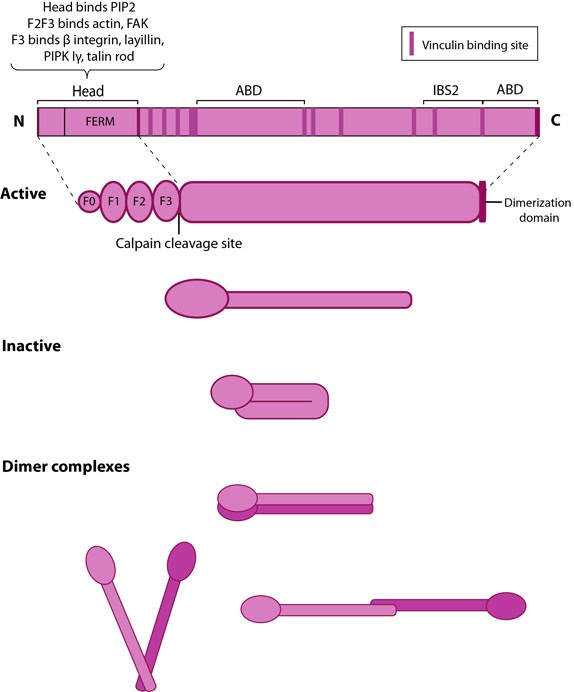
Talin structure: This schematic diagram illustrates the molecular organization of talin (reviewed in [6, 7, 18]) and shows how talin is represented throughout this resource. Hypothetical dimer complexes are presented [19]. ABD = actin binding domain, IBS2 = integrin binding site 2, PIP2 = phosphatidylinositol-4,5-bisphosphate, FAK = focal adhesion kinase, PIPK = phosphatidylinositol phosphate type 1γ.
155.
Tensin: This
schematic diagram illustrates the molecular organization of tensin and
provides examples for how tensin is represented in figures throughout
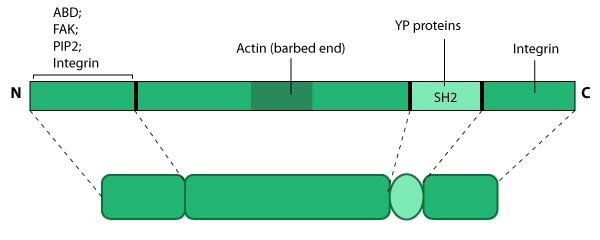
this resource. Relevant domains believed to be important for binding to actin and for protein-protein interactions are highlighted (reviewed in [7840759, 14592531]). PIP2- phosphatidylinositol [4, 5]-bis-phosphate; YP- phosphotyrosine; SH2- Src homology 2.
156.
Thermodynamic model for mechanosensing and self-assembly of focal adhesions: FAs are considered to be elastic, made of smaller units represented by the hexagonal building blocks that act like short springs. A. These blocks consisting of FA components are scattered outside the adhesion structure and when added to FA, they attach to the ECM (blue slab) through peg-like structures on the cell surface i.e., receptors. B. When FA is pulled by forces from actomyosin contractions (green arrows), the constituent aggregates get stretched and tension prevails. C. This drives the self-assembly at points of force application by insertion of new aggregates leading to energy relaxation. Adapted from [16930976, 19598236].
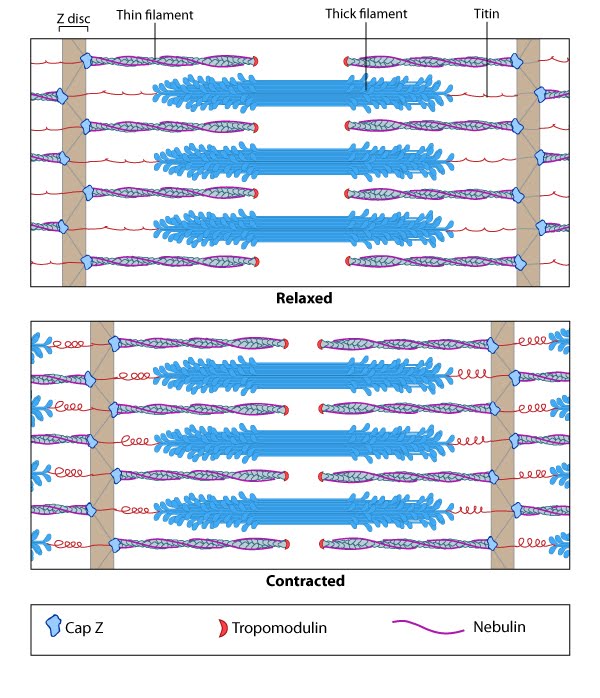
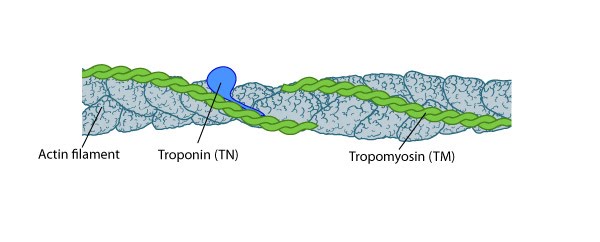
Tropomyosin stabilizes thin filaments: TM binds to the side of adjacent actin subunits along the groove of the helix to stabilize and stiffen the actin filament [4894870]. TM also prevents other proteins from accessing the filament; this inhibition is essential for regulating muscle contraction [17178912]. TN controls the positioning of TM along the groove of the actin filament.
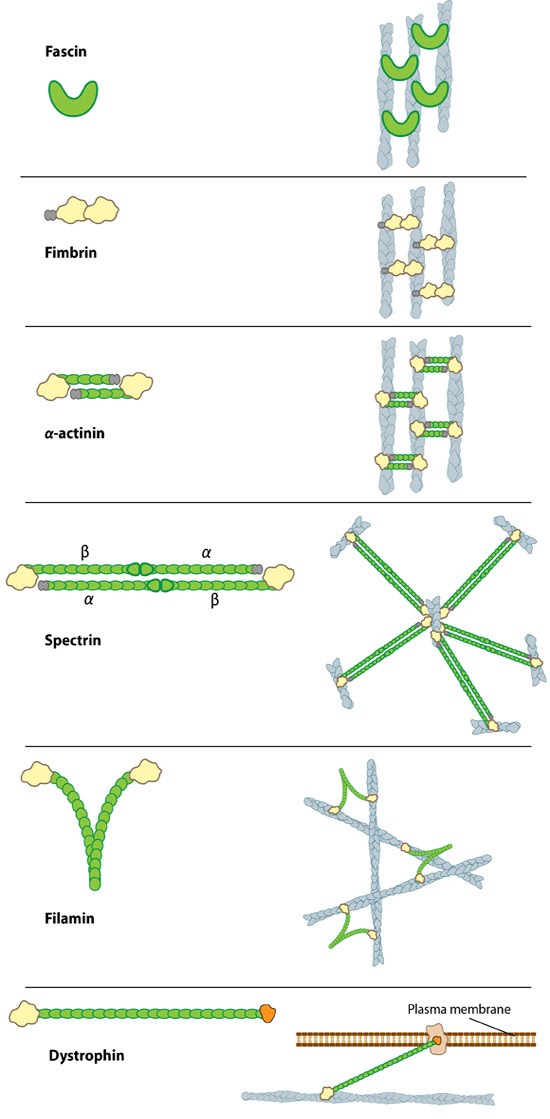
Types of actin filament crosslinking proteins: Smaller cross-linking proteins that are more globular (e.g. fascin) or have more than one actin binding site (e.g. fimbrin, α-actinin dimers) primarily form actin bundles. Larger crosslinking proteins (e.g. spectrin, filamin, dystrophin) create more space between actin filaments and they generally form actin networks. Other actin crosslinking proteins not shown here include: scruin, dematin, and villin.
161.
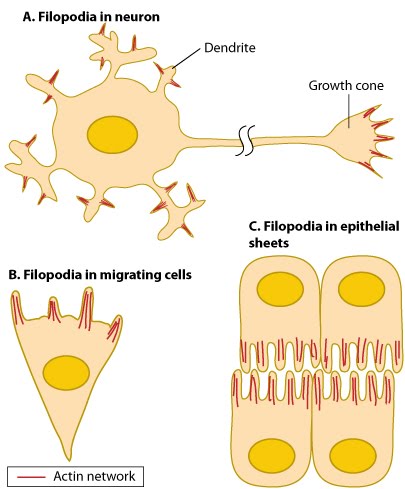
Types of adhesions found in filopodia.: Three distinct adhesion types are found in filopodia: tip adhesions serve as guidance cues, shaft adhesions help control lamellipodial advance, and basal adhesions assist with filopodia emergence and are involved with components linking the cell to the substratum.
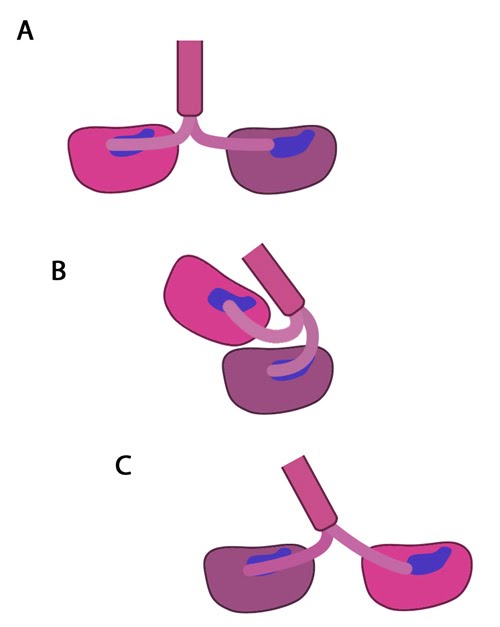
Types of cadherin interactions: All cadherins have a common extracellular domain that is structured into tandem blocks that are variable in number and are called 'cadherin repeats'; classical cadherins have five cadherin repeats (as shown). The cadherin intracellular domain is bound by adaptor proteins. (A) Example of a cis interaction (i.e. on the same cell) that is homophilic (i.e. cadherins of the same type). (B) Example of a trans interaction (i.e. between two cells) that is homophilic. (C) Example of a trans interaction that is heterophilic (between two different types of cadherin).
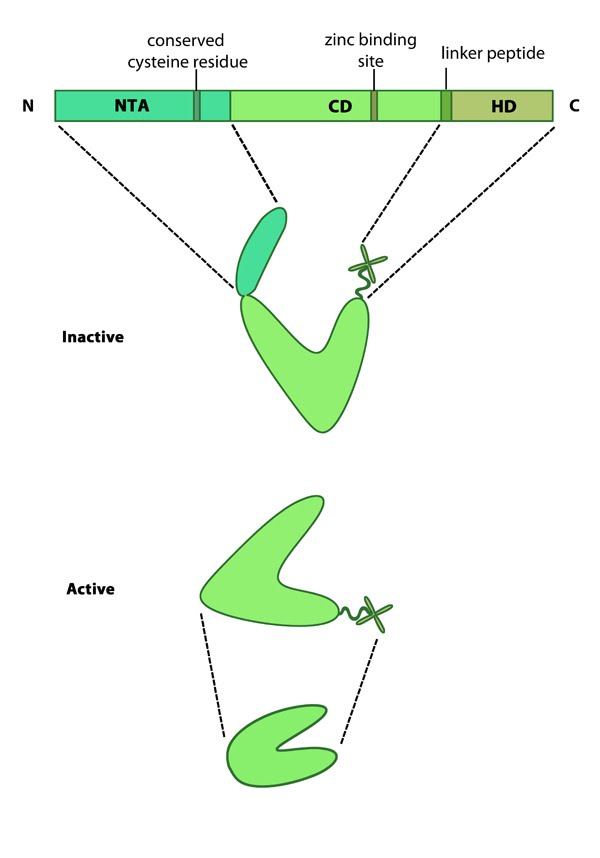
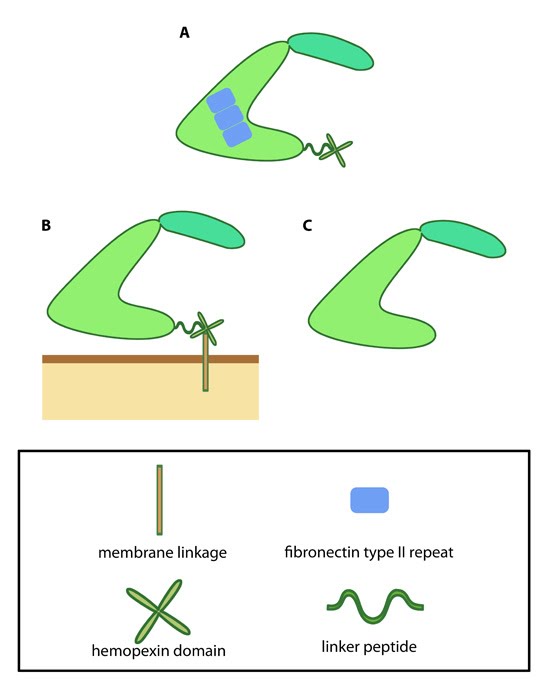
Variations in MMP structure: This schematic diagram illustrates variations on the common MMP structure (reviewed in [3]). (A) MMP2 and MMP9 have additional fibronectin domain inserts within the catalytic domain (B)
Membrane bound MMPs (MT-MMPs) contain an additional membrane linkage
domain attached to the hemopexin domain, in the form of either a GPI
(glycosylphosphatidylinositol) anchor or a transmembrane domain. (C) Minimal MMPs, such as MMP7 and MMP26, do not contain a linker peptide or hemopexin domain.
165.
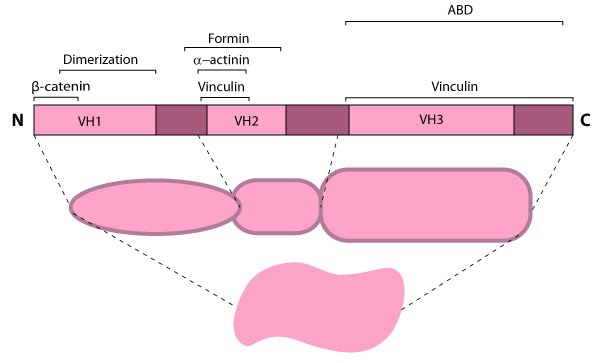
Vinculin: This schematic diagram illustrates the molecular organization of vinculin and shows how vinculin is represented throughout this resource. Relevant domains believed to be important for actin binding and protein-protein interactions are highlighted (reviewed in [16893648]).
166.
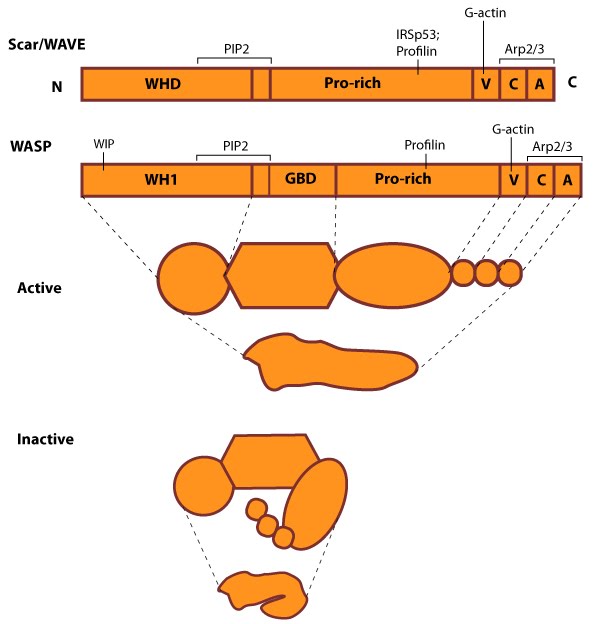
WASP_WAVE_nucleation_promoting_factors: The molecular organization of Scar/WAVE family members and WASP, with examples of how the NPFs are represented in figures throughout this resource. Relevant domains believed to be important for binding to actin and for protein-protein interactions are highlighted (reviewed in [2, 13]). WHD= WAVE (also known as SCAR)-homology domain; WIP= WASP-interacting protein; V= verprolin-homology domain (i.e. WASP-homology-2 domain [WH2]); C= cofilin-homology domain; A= acidic-rich domain; WH1= WASP-homology-1 (aka Ena-VASP-homology-1 [EVH1]); GBD= GTPase binding domain. Of note: N-WASP contains an additional V domain, and the Scar/WAVE proteins lack a direct binding domain for the Rho GTPase family.
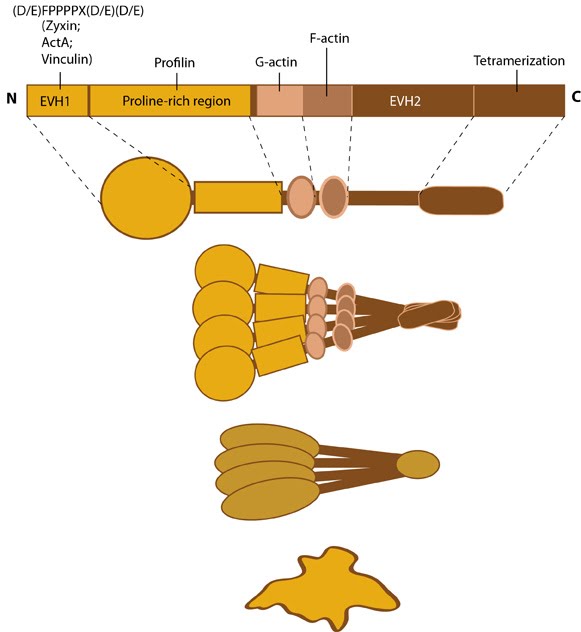
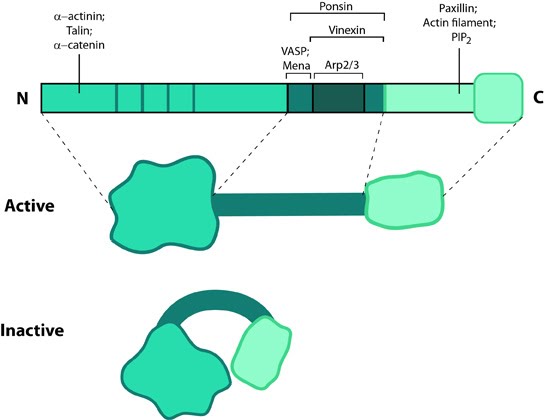
Zyxin: This schematic diagram illustrates the molecular organization of zyxin and provides examples for how zyxin is represented in figures throughout this resource.
169.
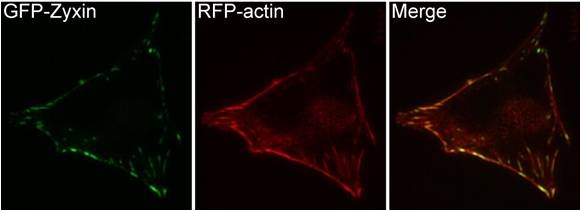
Zyxin localization: A NIH3T3 cell, plated on a collagen coated glass slide and transfected with RFP-actin and GFP-zyxin. The basal surface of the cell was imaged using TIRF microscopy, on an Olympus IX81 Inverted microscope at 60x magnification. [Image captured by Machiyama Hiroaki, Mechanobiology Institute, Singapore]
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}