Cellular Organization[Edit]
In this topic
Cells are the basic units of life; small machines that facilitate and sustain every process within a living organism. Muscle cells contract to maintain a heartbeat and allow us to move, neurons form networks that give rise to memories and enable thought processes. Epithelial cells arrange to form surface barriers between tissues and the many cavities throughout our bodies.
Not only do different cell types facilitate unique functions, but their molecular, genetic and structural compositions may also differ. For this reason, different cell types often possess variations in phenotype, such as cell size and shape.
A cell’s function is achieved through the culmination of hundreds of smaller processes, many of which are dependent on each other, and share protein or molecular components. Despite the phenotypic and functional variations that exist between cell types, it remains true that a high level of similarity exists when exploring subcellular processes, the components involved, and importantly, the organization of these components.
With most subcellular processes under precise regulatory control of other subcellular processes, and with components often shared between different molecular pathways and protein cascades, cellular organization is of great importance. This is true for every cell type, with compartmentalization of subcellular processes, and protein localization, recruitment and delivery ensuring they are constantly repeated in an efficient manner and with accurate results.
At a basic level eukaryotic cells can be described as containing three distinct sub-cellular regions; namely the membrane, the cytosol and the nucleus. However cellular compartmentalization is further complicated by the abundance of specific organelles.
Despite being just several nanometers in width, cellular membranes are highly enriched in signaling receptors, transmembrane mechanosensors, pumps and channels, and, depending on their makeup, can recruit and retain a pool of mechanosensors important in the field of mechanobiology. In many cases these mechanosensors interact with the cytoskeleton, which resides in close proximity to the membrane. The cytosol on the other hand houses cellular organelles, including the Golgi, the endoplasmic reticulum (ER), ribosomes and numerous vesicles and vacuoles. Soluble proteins may exist in this region. Meanwhile, the nucleus houses the genetic material and all components related to its expression and regulation. Although the processes of the nucleus are not as well established in terms of their role in mechanobiology, recent findings indicate several important connections, often with the mechanotransduction signaling pathways culminating in alterations to gene expression.
Each of these sub-cellular regions must work coherently for the survival and efficient functioning of the cell. The adequate organization of organelles, proteins and other molecules throughout each region allows individual protein components to function in concert with each other, effectively driving individual subcellular processes that culminate in an overall cellular function.
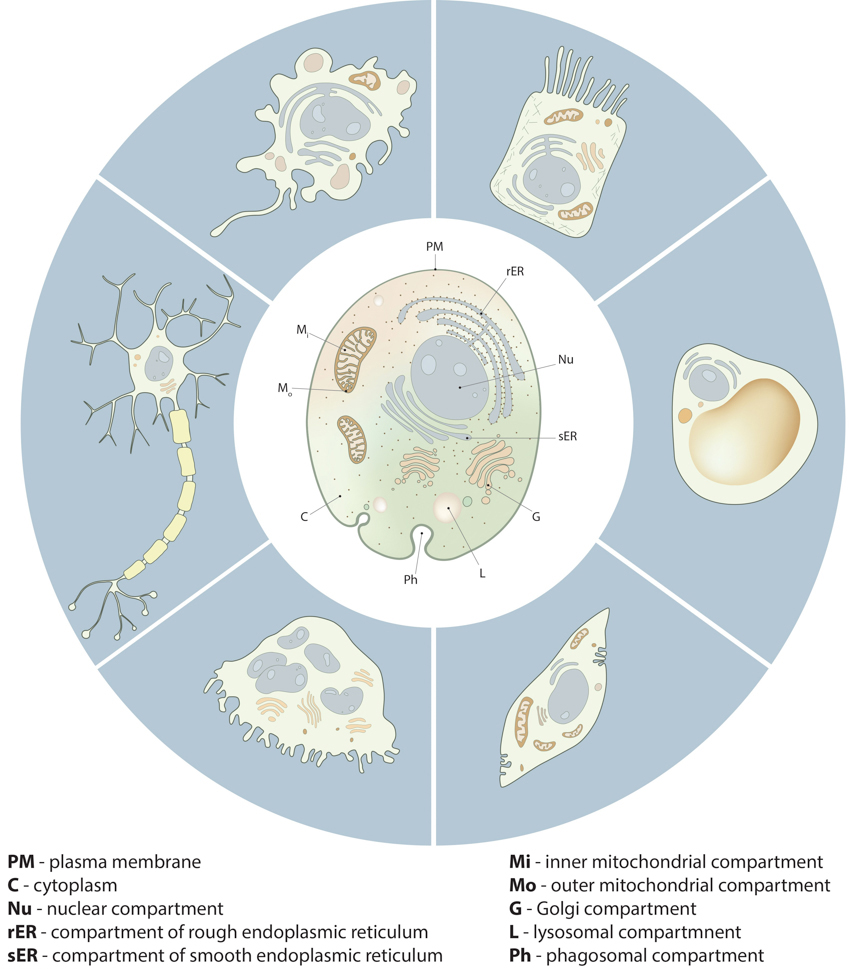 Figure 1. Compartmentalization in cells:: Despite the morphological and functional variety of cells from different tissue types and different organisms, all cells share important similarities in their compartmental organization. These fundamental compartments, often referred to as organelles, are summarized in the drawing of the generic animal cell (central cell). Examples of specialized cell types, shown around the generic cell, include neuron, macrophage, intestine epithelial cell, adipocyte, muscle cell and osteoclast.
Compartmentalization increases the efficiency of many subcellular processes by concentrating the required components to a confined space within the cell. Where a specific condition is required to facilitate a given subcellular process, this may be locally contained so as not to disrupt the function of other subcellular compartments. For example, lysosomes require a lower pH in order to facilitate degradation of internalized material. Membrane bound proton pumps present on the lysosome maintain this condition. Similarly, a large membrane surface area is required by mitochondria to efficiently generate ATP from electron gradients across its lipid bilayer. This is achieved through the structural composition of this particular organelle.
Figure 1. Compartmentalization in cells:: Despite the morphological and functional variety of cells from different tissue types and different organisms, all cells share important similarities in their compartmental organization. These fundamental compartments, often referred to as organelles, are summarized in the drawing of the generic animal cell (central cell). Examples of specialized cell types, shown around the generic cell, include neuron, macrophage, intestine epithelial cell, adipocyte, muscle cell and osteoclast.
Compartmentalization increases the efficiency of many subcellular processes by concentrating the required components to a confined space within the cell. Where a specific condition is required to facilitate a given subcellular process, this may be locally contained so as not to disrupt the function of other subcellular compartments. For example, lysosomes require a lower pH in order to facilitate degradation of internalized material. Membrane bound proton pumps present on the lysosome maintain this condition. Similarly, a large membrane surface area is required by mitochondria to efficiently generate ATP from electron gradients across its lipid bilayer. This is achieved through the structural composition of this particular organelle.
Importantly, individual organelles may be transported throughout the cell, and this essentially localizes entire subcellular processes to regions where they are required. This has been observed in neurons, which have extremely long axonal processes and require mitochondria to generate ATP at various locations along the axon. It would be inefficient to rely on the passive diffusion of ATP down the length of the axon.
Compartmentalization can also have important physiological implications. For example, polarized epithelial cells, which possess distinct apical and basolateral membranes, can, for instance produce a secretory surface for various glands. Similarly, neuronal cells develop effective networks due to the production of dendrites and axonal processes from opposite ends of the cell body. Moreover, in the case of embryonic stem cells, cell polarization can result in distinct fates of the daughter cells.
With each organelle facilitating its own function, they may be considered as subcellular compartments in their own right. However, without a regulated supply of components to the compartment, the processes and mechanisms that produce their overall function will be impeded. With many proteins and molecular components participating in multiple subcellular processes, and therefore required throughout multiple subcellular compartments, effective transport of the protein and molecular components, either by passive diffusion or directed recruitment, is essential for the overall function of the cell.
 Figure 2. An
overview of co-translational targeting of proteins destined for secretion or
membrane insertion: SRP (signal recognition particle) interacts with the signal
sequence as soon as it emerges from the ribosomal polypeptide exit tunnel (1-2).
In eukaryotes peptide elongation pauses upon SRP / ribosome nascent chain complex
formation; the complex is then targeted to the ER membrane by the interaction
with the SRP receptor (3). GTP binding to SRP and SRP receptor is a
prerequisite for SRP / SRP receptor complex formation. Following the complex
positioning (3), SRP / SRP receptor complex dissociates from the peptide due to
GTP hydrolysis. The growing peptide is gradually transferred via the
protein-conducting channel (the translocon) into the ER lumen (4). Usually, signal sequences are cleaved off co-translationally by
SPases (signal peptidases), and the resulting cleaved signal sequences are
termed signal peptides (4-5).Protein recruitment is essentially a form of protein recognition, made possible by the presence of specific amino-acid sequences within the protein structure. For example, many membrane bound proteins carry signal peptides that are recognized by signal receptors that guide them to the target site. The nuclear localization signal is one such example. Proteins that are destined for the endoplasmic reticulum also carry a signal peptide.
Figure 2. An
overview of co-translational targeting of proteins destined for secretion or
membrane insertion: SRP (signal recognition particle) interacts with the signal
sequence as soon as it emerges from the ribosomal polypeptide exit tunnel (1-2).
In eukaryotes peptide elongation pauses upon SRP / ribosome nascent chain complex
formation; the complex is then targeted to the ER membrane by the interaction
with the SRP receptor (3). GTP binding to SRP and SRP receptor is a
prerequisite for SRP / SRP receptor complex formation. Following the complex
positioning (3), SRP / SRP receptor complex dissociates from the peptide due to
GTP hydrolysis. The growing peptide is gradually transferred via the
protein-conducting channel (the translocon) into the ER lumen (4). Usually, signal sequences are cleaved off co-translationally by
SPases (signal peptidases), and the resulting cleaved signal sequences are
termed signal peptides (4-5).Protein recruitment is essentially a form of protein recognition, made possible by the presence of specific amino-acid sequences within the protein structure. For example, many membrane bound proteins carry signal peptides that are recognized by signal receptors that guide them to the target site. The nuclear localization signal is one such example. Proteins that are destined for the endoplasmic reticulum also carry a signal peptide.
In other cases proteins may carry a signal patch. This usually consists of about 30 amino acids that are not present in a linear sequence, but are in close spatial proximity in the 3-dimensional space.
Interestingly, the organization of a cell, and its various regions, do play a role in directing the recruitment of proteins to a given site. For example, in epithelial cells, which are polarized, protein composition at the apical membrane is very different from that at the basolateral membrane. This is achieved through the recognition of distinct signal sequences that target proteins to each of these regions. For example, apical membrane proteins are very often anchored to GPI (glycophosphatidylinositol) while basolateral proteins have diLeu or tyrosine amino acid based signature sequences [1].
Cytosolic proteins that are associated with the plasma membranes are often localized based on their interaction with membrane lipids such as the phosphoinositides. Phosphatidylinositol 4,5 bisphosphate (PIP2) is the most abundant phosphoinositide in mammalian cells. PIP2 is enriched in certain regions on the membrane, and due to its ability to interact with the PH (pleckstrin homology) domains of proteins; it can locally recruit proteins possessing such domains. FERM domain containing proteins, that link actin with the membrane, can also interact with PIP2 [2]. Another closely related phosphoinositide, Phosphatidylinositol 3,4,5 triphosphate (PIP3) may also be enriched in certain regions of the plasma membrane. The enrichment of PIP3 has been shown to accumulate at tips of neurons thus inducing cell polarization and axon formation [3]. It is also suspected to induce local actin depolymerization in Dictyostelium cells and influence cell migration [4] although other studies in neutrophils have indicated its role is in actin polymerization [5].
Additionally, cytosolic domains of transmembrane proteins can attract other proteins present in the cytosol that participate in signaling cascades. For example, Receptor Tyrosine Kinases (RTKs) phosphorylate certain tyrosine residues in the cytosolic domains of upon ligand binding to the extracellular domains. The phosphorylated tyrosines attract proteins possessing SH2 (src homology) or PTB (phosphotyrosine binding) domains, which comprise a signaling complex.
The curvature of membranes can also enrich proteins of certain permissible sizes and mechanical properties. For example trans-membrane proteins with conical shape such as nicotinic acetylcholine receptor prefer curved regions in the membrane as this relaxes the membrane tension and shape. Sorting of membrane constituents that prefer curvature has been observed in the Golgi cisternae rims thus reducing mechanical stress in the flatter cisternal regions [6, 7]. Proteins containing BAR (Bin/Amphiphysin/Rvs) domain can detect and induce membrane curvatures and also help sort proteins to different regions of membrane based on curvature. BAR domain proteins are particularly enriched in regions of the membrane with high endocytic activity [8, 9].
A more efficient way of maintaining the concentration of protein components is by their directed delivery via the cytoskeletal network.
The cytoskeleton, which is comprised of actin filaments and microtubules, spans the entire cell and connects the plasma membrane to the nucleus and other organelles. These filaments perform many purposes, from providing structural support to the cell, to generating the forces required for cell translocation. They may also serve as “tracks” on which motor proteins can translocate as they carry cargo from one location to another; analogous to a freight train transporting cargo along a network of railway tracks.
Delivery of components is primarily facilitated by ATP/GTP driven molecular motors such as Myosin V or Myosin X, Kinesin or Dynein. These proteins, or homologs of them, have been observed in a multitude of cell types, including yeast, plant and animal cells. The molecular motors dynein and kinesin walk on microtubules while myosin walks on actin filaments. Expectedly, these motors walk in a unidirectional manner, although not necessarily in the same direction as each other.
Microtubule based transport has been primarily studied in neuronal cells. Axons can be several microns in length (sometimes even meter-long), so there is a need to transport proteins, lipids, synaptic vesicles, mitochondria and other components along the axon. All microtubules in the axons are unidirectional, with ‘minus’ ends that point towards the cell body and ‘plus’ ends that point towards the synapse. Kinesin motors move along these tracks to ferry cargo from the cell body to the axon. Disruption of kinesin-mediated cargo transport is correlated with several neuro-muscular diseases such as spinal muscle atrophy and spinal and bulbar muscular atrophy [10, 11]. Dynein, on the other hand, plays important role in trafficking cargo in dendrites [12].
Much of our understanding on myosin-based delivery comes from studies in yeast, although mechanisms appear to be conserved up to humans. In yeast cells, myosin V motors are used to deliver vesicles to budding sites [13]. The actomyosin complex has also been implicated in the delivery of secretory vesicles to sites of growth, exocytosis and organelle segregation during cell division [14, 15, 16]. Other myosins that have been implicated in cargo transport include, myosin VI, which is important for the generation, sorting and delivery of endocytic vesicles [17], and myosin X, which carries various proteins to the tips of filopodia via their FERM domain. One example of a protein transported by Myosin X is β-integrin.
Although functionally similar, important differences exist between microtubule and actin based transport mechanisms. While microtubule based transport occurs over long distances, actin based delivery occurs over much shorter distances of 1-5 µm [18], reviewed in [19]. Furthermore, microtubules play an important role in the delivery of cargo from the ER to the Golgi and to the cell membrane [20, 21]. Actin based transport on the other hand is associated with the delivery of cargo and vesicles to the plasma membrane, with the goal of initiating rapidly growing and retracting protrusions. These protrusions sense and measure the mechanical properties of the microenvironment, facilitating cell migration.
Signaling pathways may carry a signal that originates from outside a cell or from within various compartments and usually involves the translocation of ions, solutes, proteins and secondary messengers.
All cells have surface receptors and other proteins to facilitate sensing of signals from the extracellular environment. These signals can be in the form of ions, small molecules, peptides, shear stress, mechanical forces, heat, etc. Once the signal is sensed by the surface receptor, it is transmitted to the cytoplasm usually by means of conformational change in the receptor or change in its phosphorylation status on the cytosolic side. This in turn triggers a downstream signaling cascade, which very often culminates in the nucleus. The signal usually results in change in gene expression profile of cells, assisting them to respond to the stimulus.
Subcellular compartments may also possess mechanisms that allow them to regulate each other’s functions. For instance, protein synthesis and secretory activity of the ER and the Golgi is modulated by phosphorylation of proteins or lipids on their membranes by cytosolic enzymes such as protein kinase A and protein kinase D. Calcium ions (Ca2+) are a common means of such signal transduction. Usually the concentration of these ions in the cytosol is quite low (~0.1µM), but can increase to 1µM upon induction by a signal. The ER is the storehouse of Ca2+ ions and when released in the cytosol they bind to several proteins, modulating their activity. For example, one target of Ca2+ is CaM kinase, which directly influences cell motility and migration by controlling myosin light chain kinase activity.
Not only do different cell types facilitate unique functions, but their molecular, genetic and structural compositions may also differ. For this reason, different cell types often possess variations in phenotype, such as cell size and shape.
A cell’s function is achieved through the culmination of hundreds of smaller processes, many of which are dependent on each other, and share protein or molecular components. Despite the phenotypic and functional variations that exist between cell types, it remains true that a high level of similarity exists when exploring subcellular processes, the components involved, and importantly, the organization of these components.
With most subcellular processes under precise regulatory control of other subcellular processes, and with components often shared between different molecular pathways and protein cascades, cellular organization is of great importance. This is true for every cell type, with compartmentalization of subcellular processes, and protein localization, recruitment and delivery ensuring they are constantly repeated in an efficient manner and with accurate results.
At a basic level eukaryotic cells can be described as containing three distinct sub-cellular regions; namely the membrane, the cytosol and the nucleus. However cellular compartmentalization is further complicated by the abundance of specific organelles.
Despite being just several nanometers in width, cellular membranes are highly enriched in signaling receptors, transmembrane mechanosensors, pumps and channels, and, depending on their makeup, can recruit and retain a pool of mechanosensors important in the field of mechanobiology. In many cases these mechanosensors interact with the cytoskeleton, which resides in close proximity to the membrane. The cytosol on the other hand houses cellular organelles, including the Golgi, the endoplasmic reticulum (ER), ribosomes and numerous vesicles and vacuoles. Soluble proteins may exist in this region. Meanwhile, the nucleus houses the genetic material and all components related to its expression and regulation. Although the processes of the nucleus are not as well established in terms of their role in mechanobiology, recent findings indicate several important connections, often with the mechanotransduction signaling pathways culminating in alterations to gene expression.
Each of these sub-cellular regions must work coherently for the survival and efficient functioning of the cell. The adequate organization of organelles, proteins and other molecules throughout each region allows individual protein components to function in concert with each other, effectively driving individual subcellular processes that culminate in an overall cellular function.
Compartmentalization in cells
Cells are not an amorphous mixture of proteins, lipids and other molecules. Instead, all cells are comprised of well-defined compartments, each specializing in a particular function. In many cases subcellular processes may be described based on whether they occur at the plasma membrane, within the cytosol or within membrane bound organelles such as the nucleus, Golgi apparatus or even vesicular components of the membrane trafficking system like lysosomes and endosomes.Figure 1. Compartmentalization in cells:: Despite the morphological and functional variety of cells from different tissue types and different organisms, all cells share important similarities in their compartmental organization. These fundamental compartments, often referred to as organelles, are summarized in the drawing of the generic animal cell (central cell). Examples of specialized cell types, shown around the generic cell, include neuron, macrophage, intestine epithelial cell, adipocyte, muscle cell and osteoclast.Importantly, individual organelles may be transported throughout the cell, and this essentially localizes entire subcellular processes to regions where they are required. This has been observed in neurons, which have extremely long axonal processes and require mitochondria to generate ATP at various locations along the axon. It would be inefficient to rely on the passive diffusion of ATP down the length of the axon.
Compartmentalization can also have important physiological implications. For example, polarized epithelial cells, which possess distinct apical and basolateral membranes, can, for instance produce a secretory surface for various glands. Similarly, neuronal cells develop effective networks due to the production of dendrites and axonal processes from opposite ends of the cell body. Moreover, in the case of embryonic stem cells, cell polarization can result in distinct fates of the daughter cells.
With each organelle facilitating its own function, they may be considered as subcellular compartments in their own right. However, without a regulated supply of components to the compartment, the processes and mechanisms that produce their overall function will be impeded. With many proteins and molecular components participating in multiple subcellular processes, and therefore required throughout multiple subcellular compartments, effective transport of the protein and molecular components, either by passive diffusion or directed recruitment, is essential for the overall function of the cell.
Protein Localization
In order for subcellular processes to be carried out within defined compartments or cellular regions, mechanisms must exist to ensure the required protein components are present at the sites and at an adequate concentration. The accumulation of a protein at a given site is known as protein localization.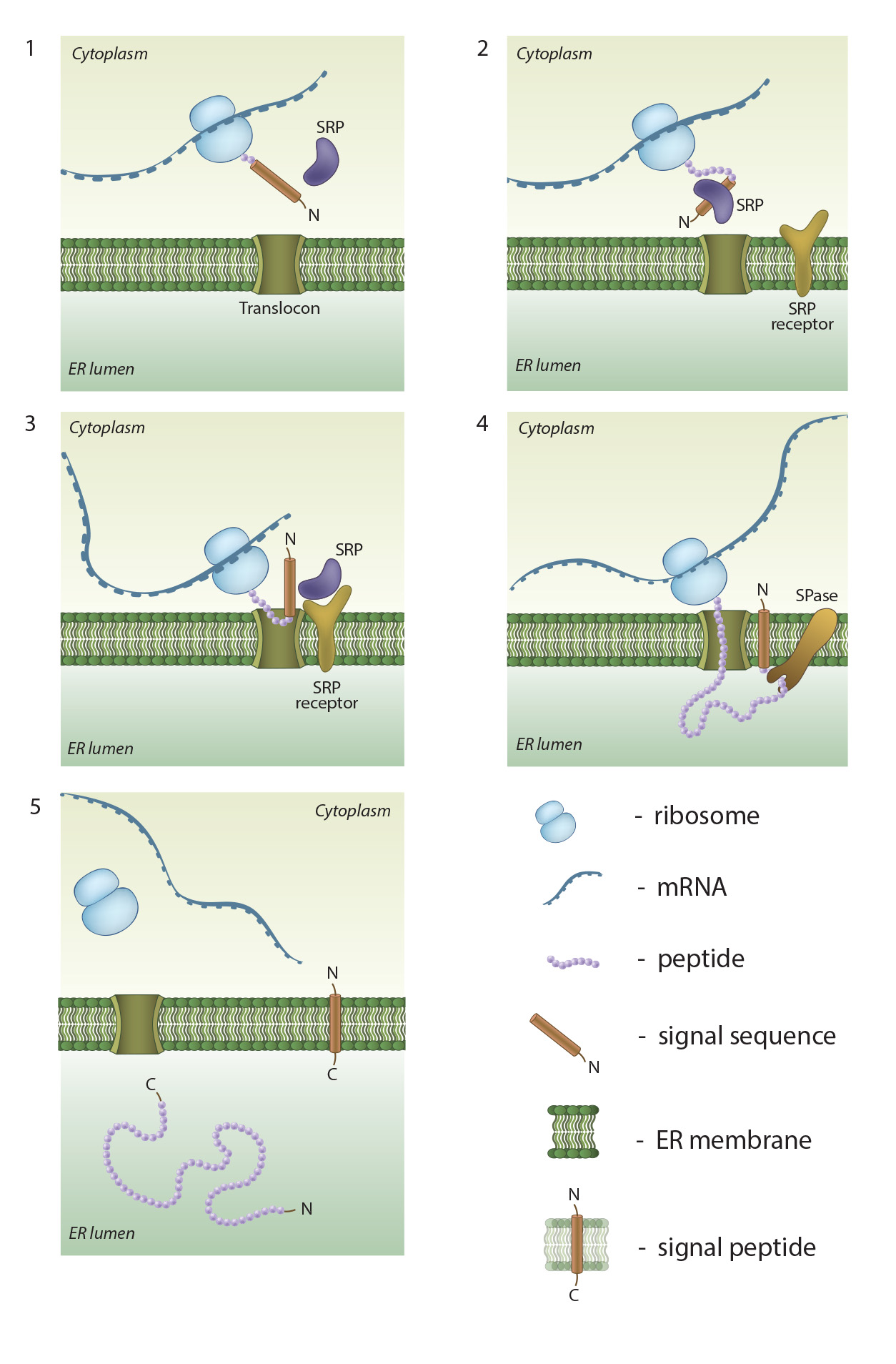 Figure 2. An
overview of co-translational targeting of proteins destined for secretion or
membrane insertion: SRP (signal recognition particle) interacts with the signal
sequence as soon as it emerges from the ribosomal polypeptide exit tunnel (1-2).
In eukaryotes peptide elongation pauses upon SRP / ribosome nascent chain complex
formation; the complex is then targeted to the ER membrane by the interaction
with the SRP receptor (3). GTP binding to SRP and SRP receptor is a
prerequisite for SRP / SRP receptor complex formation. Following the complex
positioning (3), SRP / SRP receptor complex dissociates from the peptide due to
GTP hydrolysis. The growing peptide is gradually transferred via the
protein-conducting channel (the translocon) into the ER lumen (4). Usually, signal sequences are cleaved off co-translationally by
SPases (signal peptidases), and the resulting cleaved signal sequences are
termed signal peptides (4-5).
Figure 2. An
overview of co-translational targeting of proteins destined for secretion or
membrane insertion: SRP (signal recognition particle) interacts with the signal
sequence as soon as it emerges from the ribosomal polypeptide exit tunnel (1-2).
In eukaryotes peptide elongation pauses upon SRP / ribosome nascent chain complex
formation; the complex is then targeted to the ER membrane by the interaction
with the SRP receptor (3). GTP binding to SRP and SRP receptor is a
prerequisite for SRP / SRP receptor complex formation. Following the complex
positioning (3), SRP / SRP receptor complex dissociates from the peptide due to
GTP hydrolysis. The growing peptide is gradually transferred via the
protein-conducting channel (the translocon) into the ER lumen (4). Usually, signal sequences are cleaved off co-translationally by
SPases (signal peptidases), and the resulting cleaved signal sequences are
termed signal peptides (4-5).In other cases proteins may carry a signal patch. This usually consists of about 30 amino acids that are not present in a linear sequence, but are in close spatial proximity in the 3-dimensional space.
Interestingly, the organization of a cell, and its various regions, do play a role in directing the recruitment of proteins to a given site. For example, in epithelial cells, which are polarized, protein composition at the apical membrane is very different from that at the basolateral membrane. This is achieved through the recognition of distinct signal sequences that target proteins to each of these regions. For example, apical membrane proteins are very often anchored to GPI (glycophosphatidylinositol) while basolateral proteins have diLeu or tyrosine amino acid based signature sequences [1].
Cytosolic proteins that are associated with the plasma membranes are often localized based on their interaction with membrane lipids such as the phosphoinositides. Phosphatidylinositol 4,5 bisphosphate (PIP2) is the most abundant phosphoinositide in mammalian cells. PIP2 is enriched in certain regions on the membrane, and due to its ability to interact with the PH (pleckstrin homology) domains of proteins; it can locally recruit proteins possessing such domains. FERM domain containing proteins, that link actin with the membrane, can also interact with PIP2 [2]. Another closely related phosphoinositide, Phosphatidylinositol 3,4,5 triphosphate (PIP3) may also be enriched in certain regions of the plasma membrane. The enrichment of PIP3 has been shown to accumulate at tips of neurons thus inducing cell polarization and axon formation [3]. It is also suspected to induce local actin depolymerization in Dictyostelium cells and influence cell migration [4] although other studies in neutrophils have indicated its role is in actin polymerization [5].
Additionally, cytosolic domains of transmembrane proteins can attract other proteins present in the cytosol that participate in signaling cascades. For example, Receptor Tyrosine Kinases (RTKs) phosphorylate certain tyrosine residues in the cytosolic domains of upon ligand binding to the extracellular domains. The phosphorylated tyrosines attract proteins possessing SH2 (src homology) or PTB (phosphotyrosine binding) domains, which comprise a signaling complex.
The curvature of membranes can also enrich proteins of certain permissible sizes and mechanical properties. For example trans-membrane proteins with conical shape such as nicotinic acetylcholine receptor prefer curved regions in the membrane as this relaxes the membrane tension and shape. Sorting of membrane constituents that prefer curvature has been observed in the Golgi cisternae rims thus reducing mechanical stress in the flatter cisternal regions [6, 7]. Proteins containing BAR (Bin/Amphiphysin/Rvs) domain can detect and induce membrane curvatures and also help sort proteins to different regions of membrane based on curvature. BAR domain proteins are particularly enriched in regions of the membrane with high endocytic activity [8, 9].
Directed Delivery of Components
Protein localization can result from the recognition of passively diffusing soluble proteins or protein complexes; however, this may not guarantee a sufficient concentration of components to maintain a given process. This can impede its completion, particularly when carried out in regions with a limited cytoplasmic volume, such as the tip of a filopodia, or when components are rapidly turned over.A more efficient way of maintaining the concentration of protein components is by their directed delivery via the cytoskeletal network.
The cytoskeleton, which is comprised of actin filaments and microtubules, spans the entire cell and connects the plasma membrane to the nucleus and other organelles. These filaments perform many purposes, from providing structural support to the cell, to generating the forces required for cell translocation. They may also serve as “tracks” on which motor proteins can translocate as they carry cargo from one location to another; analogous to a freight train transporting cargo along a network of railway tracks.
Delivery of components is primarily facilitated by ATP/GTP driven molecular motors such as Myosin V or Myosin X, Kinesin or Dynein. These proteins, or homologs of them, have been observed in a multitude of cell types, including yeast, plant and animal cells. The molecular motors dynein and kinesin walk on microtubules while myosin walks on actin filaments. Expectedly, these motors walk in a unidirectional manner, although not necessarily in the same direction as each other.
Microtubule based transport has been primarily studied in neuronal cells. Axons can be several microns in length (sometimes even meter-long), so there is a need to transport proteins, lipids, synaptic vesicles, mitochondria and other components along the axon. All microtubules in the axons are unidirectional, with ‘minus’ ends that point towards the cell body and ‘plus’ ends that point towards the synapse. Kinesin motors move along these tracks to ferry cargo from the cell body to the axon. Disruption of kinesin-mediated cargo transport is correlated with several neuro-muscular diseases such as spinal muscle atrophy and spinal and bulbar muscular atrophy [10, 11]. Dynein, on the other hand, plays important role in trafficking cargo in dendrites [12].
Much of our understanding on myosin-based delivery comes from studies in yeast, although mechanisms appear to be conserved up to humans. In yeast cells, myosin V motors are used to deliver vesicles to budding sites [13]. The actomyosin complex has also been implicated in the delivery of secretory vesicles to sites of growth, exocytosis and organelle segregation during cell division [14, 15, 16]. Other myosins that have been implicated in cargo transport include, myosin VI, which is important for the generation, sorting and delivery of endocytic vesicles [17], and myosin X, which carries various proteins to the tips of filopodia via their FERM domain. One example of a protein transported by Myosin X is β-integrin.
Although functionally similar, important differences exist between microtubule and actin based transport mechanisms. While microtubule based transport occurs over long distances, actin based delivery occurs over much shorter distances of 1-5 µm [18], reviewed in [19]. Furthermore, microtubules play an important role in the delivery of cargo from the ER to the Golgi and to the cell membrane [20, 21]. Actin based transport on the other hand is associated with the delivery of cargo and vesicles to the plasma membrane, with the goal of initiating rapidly growing and retracting protrusions. These protrusions sense and measure the mechanical properties of the microenvironment, facilitating cell migration.
Communication Pathways
With different processes being carried out in separate subcellular compartments, organized across different regions of the cell, intracellular communication is paramount. Such communication, which is described in greater detail under “cell signaling” allows cells to maintain the concentration of specific proteins and within the correct regions, depending on the requirements of a given process or cell state. This ultimately ensures individual compartments function efficiently, and enables one subcellular process to drive another. This ultimately allows a cell to facilitate its primary functions in an efficient and coherent manner.Signaling pathways may carry a signal that originates from outside a cell or from within various compartments and usually involves the translocation of ions, solutes, proteins and secondary messengers.
All cells have surface receptors and other proteins to facilitate sensing of signals from the extracellular environment. These signals can be in the form of ions, small molecules, peptides, shear stress, mechanical forces, heat, etc. Once the signal is sensed by the surface receptor, it is transmitted to the cytoplasm usually by means of conformational change in the receptor or change in its phosphorylation status on the cytosolic side. This in turn triggers a downstream signaling cascade, which very often culminates in the nucleus. The signal usually results in change in gene expression profile of cells, assisting them to respond to the stimulus.
Subcellular compartments may also possess mechanisms that allow them to regulate each other’s functions. For instance, protein synthesis and secretory activity of the ER and the Golgi is modulated by phosphorylation of proteins or lipids on their membranes by cytosolic enzymes such as protein kinase A and protein kinase D. Calcium ions (Ca2+) are a common means of such signal transduction. Usually the concentration of these ions in the cytosol is quite low (~0.1µM), but can increase to 1µM upon induction by a signal. The ER is the storehouse of Ca2+ ions and when released in the cytosol they bind to several proteins, modulating their activity. For example, one target of Ca2+ is CaM kinase, which directly influences cell motility and migration by controlling myosin light chain kinase activity.