Cell-cell signaling[Edit]
Cell-cell signaling refers to inter-cellular communication through the transduction of chemical, mechanical or electrical signals, facilitated by the formation of specialized cell-cell adhesion junctions. There are three main types of cell-cell adhesive junctions in mammals that detect and transduce signals from neighbouring cells [1]:
Tight junctions
Also known as zonula occludens, these junctions are found in epithelial and endothelial cells and primarily function as diffusion barriers. Integral membrane proteins commonly associated with tight junctions include; claudins, occludin, tricellulin and junctional adhesion molecules (JAMs). These proteins facilitate the formation of the anastomosing membrane strands that comprise tight junctions [2].
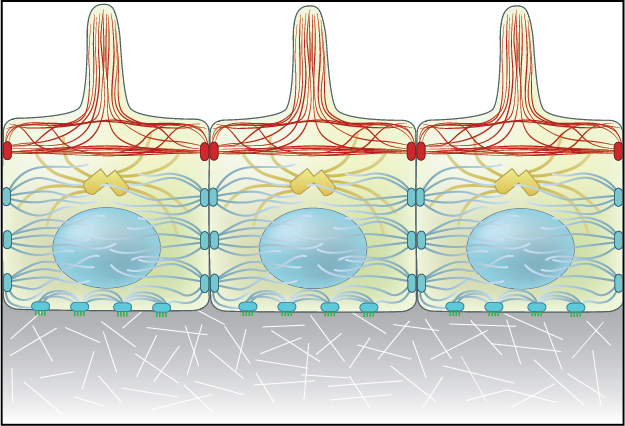 Figure 1. Adherens junctions link actin filaments between cells.: Adherens junctions link actin filaments between cells. This diagram specifically depicts adherens junctions (red rectangles) connecting actin filaments (red lines) across polarized epithelial cells. In these cells, this results in the formation of contractile bundles of actin and myosin filaments near the apical surface – this forms a structure called the adhesion belt (arrows). Other junctions termed desmosomes (large blue rectangles) and hemi-desmosomes (small blue rectangles) link intermediate filaments (blue lines) between cells.
Adherens junctions regulate cell shape, maintain tissue integrity and translate actomyosin-generated forces throughout a tissue [3]. A key component of adherens junctions are the transmembrane glycoproteins cadherins, which bind the intracellular proteins p120-catenin and β-catenin, after which α-catenin is recruited by β-catenin. A vast network of adhesion receptors, scaffolding proteins, actin regulators and signaling proteins regulate the formation of this complex, via a complex network of interactions, which is currently being characterized and has been named the cadhesome [personal communication A/P Ronen Zaidel-bar, MBI, NUS].
Figure 1. Adherens junctions link actin filaments between cells.: Adherens junctions link actin filaments between cells. This diagram specifically depicts adherens junctions (red rectangles) connecting actin filaments (red lines) across polarized epithelial cells. In these cells, this results in the formation of contractile bundles of actin and myosin filaments near the apical surface – this forms a structure called the adhesion belt (arrows). Other junctions termed desmosomes (large blue rectangles) and hemi-desmosomes (small blue rectangles) link intermediate filaments (blue lines) between cells.
Adherens junctions regulate cell shape, maintain tissue integrity and translate actomyosin-generated forces throughout a tissue [3]. A key component of adherens junctions are the transmembrane glycoproteins cadherins, which bind the intracellular proteins p120-catenin and β-catenin, after which α-catenin is recruited by β-catenin. A vast network of adhesion receptors, scaffolding proteins, actin regulators and signaling proteins regulate the formation of this complex, via a complex network of interactions, which is currently being characterized and has been named the cadhesome [personal communication A/P Ronen Zaidel-bar, MBI, NUS].
The Adherens Junction is a type of stable cell contact, or anchoring junction, that keeps cells in a tissue together by forming an interconnected lateral bridge that links the actin cytoskeleton of neighboring cells (reviewed in [4, 5]). Adherens junctions come in many forms: In cardiac cells, contractile bundles are anchored by adherens junctions [6], while in nonepithelial tissues, adherens junctions link cortical actin filaments between cells (see “cell cortex”).
In polarized epithelial cells, the most obvious feature of the actin cytoskeleton is the contractile bundle of actin filaments usually located near the apical surface; this circumferential belt (aka adhesion belt) is linked from one cell to another through the extracellular domains of a continuous band of transmembrane molecules belonging to the cadherin family [7, 8], along with other anchoring proteins (e.g. catenins, vinculin, and α-actinin) [9, 10]. The actin and associated myosin are positioned to sense tension and to brace the cell [11, 12]. Contraction of the actin network also provides motile forces for tissue morphogenesis and cell migration [13, 14].
In vertebrate epithelia, all three of the aforementioned junctions have defined spatial organizations, with tight junctions located most apically, followed by adherens junctions and lastly desmosomes [2]. Each type of junction has a distinct ultrastructure, which can additionally vary in appearance dependent on cell type and morphology.
Adherens junctions
Figure 1. Adherens junctions link actin filaments between cells.: Adherens junctions link actin filaments between cells. This diagram specifically depicts adherens junctions (red rectangles) connecting actin filaments (red lines) across polarized epithelial cells. In these cells, this results in the formation of contractile bundles of actin and myosin filaments near the apical surface – this forms a structure called the adhesion belt (arrows). Other junctions termed desmosomes (large blue rectangles) and hemi-desmosomes (small blue rectangles) link intermediate filaments (blue lines) between cells.The Adherens Junction is a type of stable cell contact, or anchoring junction, that keeps cells in a tissue together by forming an interconnected lateral bridge that links the actin cytoskeleton of neighboring cells (reviewed in [4, 5]). Adherens junctions come in many forms: In cardiac cells, contractile bundles are anchored by adherens junctions [6], while in nonepithelial tissues, adherens junctions link cortical actin filaments between cells (see “cell cortex”).
In polarized epithelial cells, the most obvious feature of the actin cytoskeleton is the contractile bundle of actin filaments usually located near the apical surface; this circumferential belt (aka adhesion belt) is linked from one cell to another through the extracellular domains of a continuous band of transmembrane molecules belonging to the cadherin family [7, 8], along with other anchoring proteins (e.g. catenins, vinculin, and α-actinin) [9, 10]. The actin and associated myosin are positioned to sense tension and to brace the cell [11, 12]. Contraction of the actin network also provides motile forces for tissue morphogenesis and cell migration [13, 14].
Desmosomes
These junctions link intermediate filaments (IFs) to the plasma membrane and in doing so provide mechanical stability to cells, which is particularly important for tissues and organs under high mechanical stress, such as the myocardium, skin and bladder. Desmosomes comprise desmosomal cadherins (desmogleins and desmocollins) that facilitate linkage between apposing cells via their extracellular domains and bind other desmosomal components intracellularly via their cytoplasmic tails. Cytoplasmic desmosomal components include plakoglobin and plakophilins, which in turn bind intermediate filaments via desmoplakin [15].In vertebrate epithelia, all three of the aforementioned junctions have defined spatial organizations, with tight junctions located most apically, followed by adherens junctions and lastly desmosomes [2]. Each type of junction has a distinct ultrastructure, which can additionally vary in appearance dependent on cell type and morphology.