Matrix metalloproteinases (MMPs) facilitate extracellular matrix disassembly[Edit]
Extracellular matrix disassembly is the degradation of the matrix components by proteases such as matrix metalloproteinases, and is a key step in normal physiological functions such as embryonic development and tissue remodeling, as well as in pathological conditions such as cancer progression.
 Figure 1. Matrix metalloproteinases (MMPs): This schematic diagram illustrates the molecular organization of MMPs (reviewed in [1]). Relevant domains believed to be important for MMP activity are highlighted (reviewed in [1]).
Mammalian MMPs comprise three major domains; an N terminal auto-inhibitory pro-domain, a catalytic domain and a C terminal hemopexin domain (see Figure at top right) [3]. MMPs are produced as inactive precursors. Their activity is inhibited through a conserved cysteine residue residing within the N terminal auto-inhibitory pro-domain. This residue interacts with the zinc ion bound to the catalytic domain and prevents cleavage of the substrate. The catalytic domain is connected to the hemopexin domain via a flexible linker peptide. The hemopexin domain forms a beta propeller structure, made of four ‘blades’ each of which represent a single hemopexin module. This domain regulates protein-protein interactions, substrate specificity and enzyme activity via interactions with tissue inhibitors of matrix metalloproteinases (TIMPs) [4].
Figure 1. Matrix metalloproteinases (MMPs): This schematic diagram illustrates the molecular organization of MMPs (reviewed in [1]). Relevant domains believed to be important for MMP activity are highlighted (reviewed in [1]).
Mammalian MMPs comprise three major domains; an N terminal auto-inhibitory pro-domain, a catalytic domain and a C terminal hemopexin domain (see Figure at top right) [3]. MMPs are produced as inactive precursors. Their activity is inhibited through a conserved cysteine residue residing within the N terminal auto-inhibitory pro-domain. This residue interacts with the zinc ion bound to the catalytic domain and prevents cleavage of the substrate. The catalytic domain is connected to the hemopexin domain via a flexible linker peptide. The hemopexin domain forms a beta propeller structure, made of four ‘blades’ each of which represent a single hemopexin module. This domain regulates protein-protein interactions, substrate specificity and enzyme activity via interactions with tissue inhibitors of matrix metalloproteinases (TIMPs) [4].
Variations in MMP structure exist (see Figure: Variations in MMP structure at bottom), with the addition or loss of certain domains (as reviewed in [3]). For example; MMP7 and MMP26 lack the hemopexin domain. MMP2 and MMP9 have additional fibronectin domain inserts within the catalytic domain. Membrane bound MMPs have additional membrane linkage domains, such as a GPI (glycosylphosphatidylinositol) anchor in the case of MMPs 14, 15, 16 and 24.
Matrix metalloproteinases (MMPs)
Matrix metalloproteinases (MMPs) are a family of proteases that digest components of the extracellular matrix (ECM) and surface receptors. The first MMP was identified in the 1960s as an enzyme which could digest the ECM component collagen and so was named collagenase [1]. Following this initial discovery a whole family of structurally related proteases were discovered. These proteases were all able to digest ECM components and did so in a zinc dependent manner. This led to the renaming of this protein family to matrix metalloproteinases. There are currently 23 human MMPs and 24 mouse MMPs [2], in addition to MMP homologs in invertebrates such as the fruit fly Drosophila melanogaster [2].MMP Structure
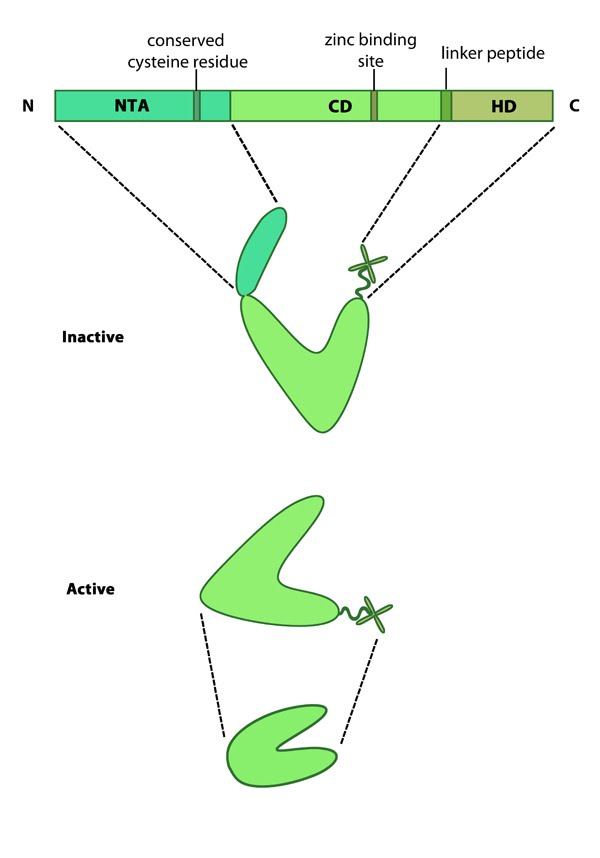 Figure 1. Matrix metalloproteinases (MMPs): This schematic diagram illustrates the molecular organization of MMPs (reviewed in [1]). Relevant domains believed to be important for MMP activity are highlighted (reviewed in [1]).
Figure 1. Matrix metalloproteinases (MMPs): This schematic diagram illustrates the molecular organization of MMPs (reviewed in [1]). Relevant domains believed to be important for MMP activity are highlighted (reviewed in [1]).Variations in MMP structure exist (see Figure: Variations in MMP structure at bottom), with the addition or loss of certain domains (as reviewed in [3]). For example; MMP7 and MMP26 lack the hemopexin domain. MMP2 and MMP9 have additional fibronectin domain inserts within the catalytic domain. Membrane bound MMPs have additional membrane linkage domains, such as a GPI (glycosylphosphatidylinositol) anchor in the case of MMPs 14, 15, 16 and 24.