
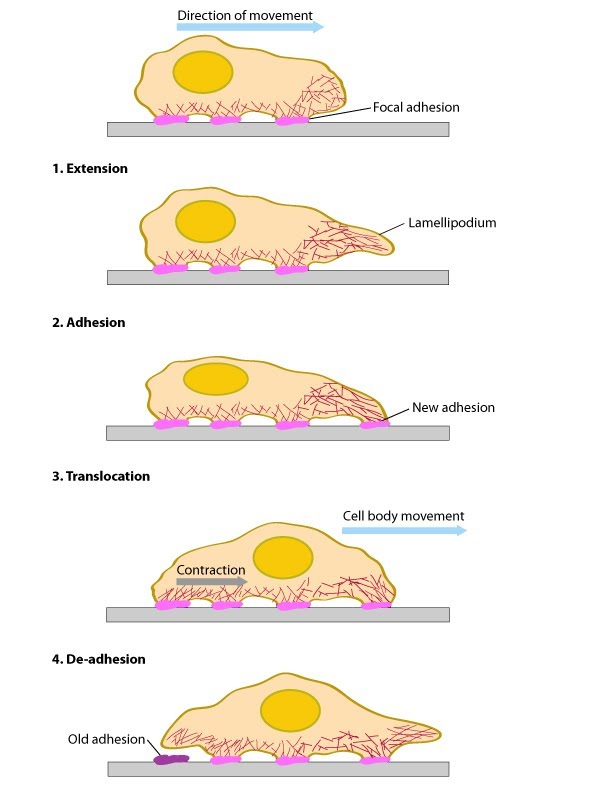
Steps in Formation 1. Initiation and Nucleation 2. Extension, Pause and Stasis 3. Formation of Adhesions 4. Force Generation and Translocation 5. Disassembly of actin filaments and retraction of the trailing edge Functional Modules
| Lamellipodia and Lamella
Steps in Formation: Overview
|
|
|
|
|
MBInfo > Cellular Structures in Mechanosensing and Cell Motility > Lamellipodia and Lamella >
Lamellipodia-Steps in formation: Overview |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites