
Functional Modules
Test Your Knowledge | 1.5 Formation of AdhesionsAs the lamellipodia continues to spread and expand forward, adhesion sites form along the leading edge (see video below). These sites not only provide traction for the cells forward movement, but permit the cell to sense and measure the rigidity of its substrate. Formation of these adhesions is a complex process, involving a number of steps and functional modules.To read more about adhesion formation please follow the link to Focal Adhesion Initiation.
Video: Focal adhesions are essential for cell spreading.
Upon neuregulin treatment, cell-surface receptors Erb B3/B4 induce
lamellipodia formation, most probably through activation of the
Rac-WAVE-Arp2/3 pathway. During lamellipodial protrusion, numerous focal
adhesions form along the cell periphery and can be visualized as
fluorescent spots (GFP-VASP). [Source: Leticia Carramusa, Weizmann
Institute of Science, Israel. Permission: Alexander Bershadsky,
Mechanobiology Institute, Singapore.]
Particularly important in lamellipodia function is the spatial arrangement of adhesions at the leading edge. Lamellipodia extension and retraction is not uniform across the whole cell and similarly, adhesion sites will not be uniformly distributed. Instead a number of factors may influence arrangement including chemistry of the extracellular matrix, substrate stiffness and cells growth conditions, as described under ‘matrix properties’. Interestingly, distribution of adhesion sites was found to be coupled to actin polymerization, with conformationally activated, but unliganded β1 integrins, shown to be interacting with elongating filaments [1]. Importantly, this interaction permits a sideways distribution along the leading edge, ensuring clusters of β1 integrin are positioned at the very front of cell protrusions. β1 integrin was also found, in the same study, to localize at the tips of growth cone filopodia [1]. Latest Findings Paxillin, a scaffold protein, has recently been shown to integrate physical cues sensed by adhesion complexes in order to confer directionality for lamellipodial protrusions [2]. Focal Adhesions as Molecular Clutches:Once formed, focal adhesions essentially act as “molecular clutches”, promoting protrusion at the leading edge whilst suppressing membrane contraction (reviewed in [3, 4, 5]). Adhesions aid forward movement by regulating the forces produced by actin dynamics in different cellular compartments through several methods:1) They aid membrane protrusion by resisting actin retrograde flow [6] and hence, indirectly promote the force produced by lamellipodial actin polymerization. 2) They convert myosin pulling forces at the lamellar interface into traction forces against the ECM that pulls the cell body forward [7, 8]. These two methods are interdependent and cooperatively contribute to the propulsive forces generated at the leading edge. The efficiency of the molecular clutch in converting this force into protrusion is variable. Because some components of the adhesion complex move along with the retrograde flow [9], the clutch slips [10]. The rate of forward protrusion increases however when actin and actin-adhesion linking components become more organized [11] and the retrograde flow of actin is subsequently slowed down [6, 12]. In this case the clutch can be described as “partially engaged”. As the adhesions grow and mature under stress, the clutch transforms from being “partially/locally engaged” to “engaged” [3] and hence can influence global cell behavior [13, 14]. Experiments have demonstrated that a biphasic relationship exists between the rate of actin flow and traction stress [15]. Whilst they are inversely related in the lamellipodium where nascent adhesions are formed and actin flow rate is high, the relationship becomes linear in areas with larger adhesions and slow actin flow [6], generating maximal propulsion at intermediate flow rates [16]. Two recent studies propose stochastic models that explain the state of adhesion clutches in these two regimes [17, 18] (reviewed in [19]). One study describes how adhesion stability is modulated by the competition between energy required for maintaining the elastic bonds at the moving actin-clutch interface, and the energy dissipation that occurs when myosin pulls in the viscoelastic actin interior [17]. When the transmitted force and the speed of actin flow are moderate, much of the energy is invested in producing traction on the substrate. At this critical transition range, either an increase or decrease in the ratio of bound adhesion complexes could theoretically occur. In the second study however, based on known experimental results, it was assumed that at moderate actin flow rates adhesion clutches reach a quasi-equilibrium state between bound and unbound forms [18]. Thus the catch bond model of receptor-actin interactions are more convincing than the slip bond since it is more likely to lead to clutch engagement and adhesion growth, as observed in the experimental studies. To find out more on the forces generated by the within the lamellipodia, read Step 4: Force Generation and Translocation. 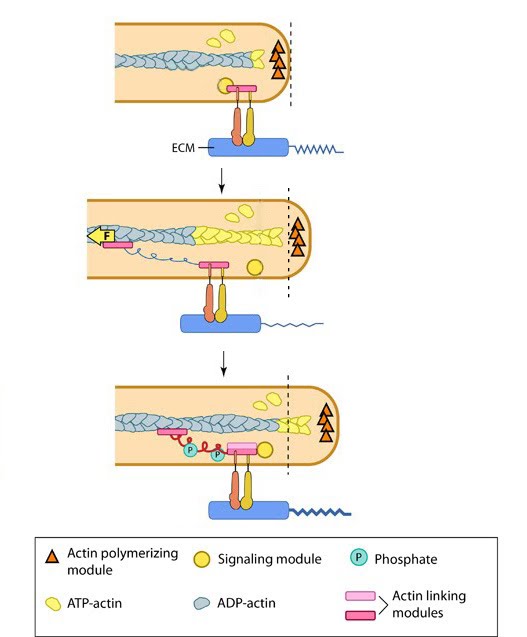 |
|
|
|
Current MCMF Content |
MBInfo > Topics > Cellular Structures in Mechanosensing and Cell Motility > Lamellipodia and Lamella > Steps in the Formation and Function of Lamella and Lamellipodia >
Adhesion Formation in Lamellipodia and Lamella |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites