
Steps in Formation 1. Initiation 2. Extension 3. Lateral movement and Stasis 4. Adherence 5. Pulling 6. Retraction and Collapse Functional Modules
| FilopodiaStep 2. Cross-linking and ExtensionProtrusion from cells occurs by a treadmilling mechanism that is common to many actin based structures [1]. According to this model, actin filaments elongate at their barbed ends at the tips of filopodia and release subunits from their pointed ends, which lie at the rear of the structure. Filopodia may assemble in a process termed “convergent elongation” from Arp2/3-nucleated lamellipodial actin filaments that have coalesced together to form parallel bundles. Under these circumstances, the barbed ends of the filaments are locally associated with each other and protected from capping [2]. This protection is provided by the activity of additional proteins, such as the Ena/VASP family of proteins, which subsequently enhance filament elongation and promote F-actin bundling thereby stimulating filopodial protrusion [2, 3, 4, 5, 6]. The bundling protein, fascin, is also enriched near the tip of the bundle and contributes to this process. Actin filaments in filopodia are unbranched [7], suggesting that assembly occurs by elongation and not by branched nucleation.Table: Filopodial Extension Rates
Protrusion of filopodia, produced by either convergent elongation or formin-nucleated filaments, must overcome the membrane curvature rigidity. This is aided by crosslinking actin filaments, which gives the structure the strength required to push against the compressive force of the plasma membrane [6, 15]. In nerve growth cones, more than 15 parallel filaments may be bundled together by crosslinking [16]. Bundle stiffness increases with the number of bundled filaments and so contributes to the overall length of the filopodium [6]. Many F-actin crosslinking proteins, such as α-actinin and fascin, co-localize at the base of filopodia and work in concert to produce crosslinked filaments [17]. The filamin family of proteins are also crucial actin crosslinking and scaffolding proteins and bind to both actin and a number of signaling molecules, including Rho GTPases. Crosslinking also increases the ATPase activity of myosins and increases the tension on filaments [18]. More on cross linking can be found in the Functional Module: Cross-linkers in Actin Bundling. 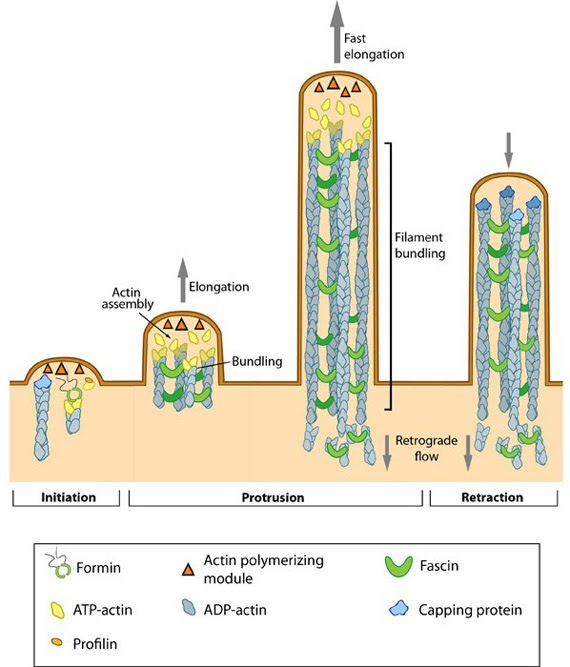 Figure: Steps in filopodium formation. Actin filament assembly can be initiated by uncapping pre-formed actin filaments or by de novo formation (which includes both formin- and Arp2/3-mediated [not shown] nucleation). The force produced by actin assembly at the barbed end of actin filaments drives membrane protrusion. Numerous proteins (including IRSp53, Ena/VASP proteins, WASp/Scar proteins) cooperate to promote actin-assembly and enhance bundling of actin filaments by fascin. When the barbed end of the filament is capped, this stops filament assembly and protrusion; continued retrograde movement of the filaments results in retraction of the filopodium. (Figure adapted from [19]). The extension rate of a filopodium is controlled by the availability of G-actin-ATP, associated structural components and the energetics of membrane bending. The growth of long filopodia (>10 μm in length) requires the rapid transport of key materials towards the growing end [20] and this process is facilitated by the myosin X motor protein using an ATP-dependent ‘walking’ mechanism. More on this process can be found in the Functional Module: Myosin-X and Cargo Transport. |
|
|
|
|
MBInfo > Cellular Structures in Mechanosensing and Cell Motility > Filopodia > Steps in Filopodia Formation and Movement >
2. Cross-linking and Extension |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites