
Steps in Formation 1. Initiation 2. Extension 3. Lateral movement and Stasis 4. Adherence 5. Pulling 6. Retraction and Collapse Functional Modules
|
|
|
|
|
|
MBInfo > Cellular Structures in Mechanosensing and Cell Motility > Filopodia > Steps in Filopodia Formation and Movement >
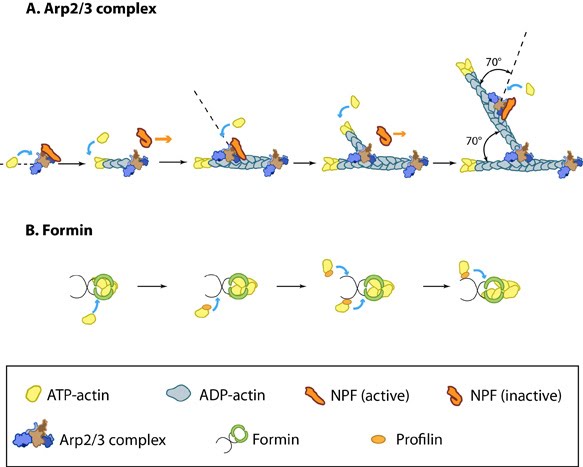
1. Initiation and Nucleation |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites