
Steps in Formation 1. Initiation 2. Extension 3. Lateral movement and Stasis 4. Adherence 5. Pulling 6. Retraction and Collapse Functional Modules | FilopodiaStep 5. PullingAlthough a reterograde motion of actin filaments is intrinsic in the formation of filopodia, the forces generated by actin treadmilling are too weak to facilitate the “pulling” mechanism required for rigidity sensing and other mechanosensing processes. This characteristic of filopodia is instead produced by the activity of myosin motor proteins such as Myosin II [1]. This process, as described in the Functional Module: ‘Myosin II in Filament Retraction’, is similar to that observed in the actin filament networks of the lamellipoda and lamella. Disassembly of actin bundles at the filopodial tip combined with the membrane tension of the lipid bilayer also theoretically supports the forces needed for pulling [2].Filopodia pulling is important in a number of processes additional to mechanosensing. One example is in the immunological response to pathogens where immune cells such as macrophages will pull a pathogen toward the cell body for active uptake and processing [3]. In another example the filopodia of mature osteoclasts bind to extracellular substrates at their tips and transport the particles rearward by retraction for resorption to the cell body, leaving the cell body adhered to the substrate [4]. It has also been shown that matrix coated beads are pulled rearward by an active process rather than by diffusion after the beads bind to integrins [1, 5]. In each case the mechanism behind the pulling remains the same. The powerstroke of Myosin II is essentially the driving force behind the movement of filaments, however, using the pulling of pathogens to macrophage cell bodies as an example it has been shown that this process occurs in three distinct stages, each defined by varying rates of retraction. Initially, and immediately following pathogen binding, a slow motion retraction of the filopodia commences. This is followed by a rapid movement of the pathogen towards the cell which is subsequently followed by another slow retraction which ends with the pathogen being fixed to the cell surface (see Figure, right panel). This final phase of retraction is also characterized by a thickening of the filopodial base which is necessary to build-up large retraction forces for internalization [2, 4]. Filopodial attachment to a surface (in addition to the ligand) produces counteracting adhesion forces which influence the retraction speed. In general, the rearward movement or ‘step size’ of a retracting filopodia decreases as the filopodia encounters a greater counteracting force [1]. Importantly, the mechanism may also be hijacked by a pathogen to ensure an efficient infection. This has been described for the Murine Leukemia Virus, amongst others [2]. In this case the virus binds to surface receptors at the tips of the filopodia and essentially rides the retracting actin filaments to entry points in the cell where it is internalized. 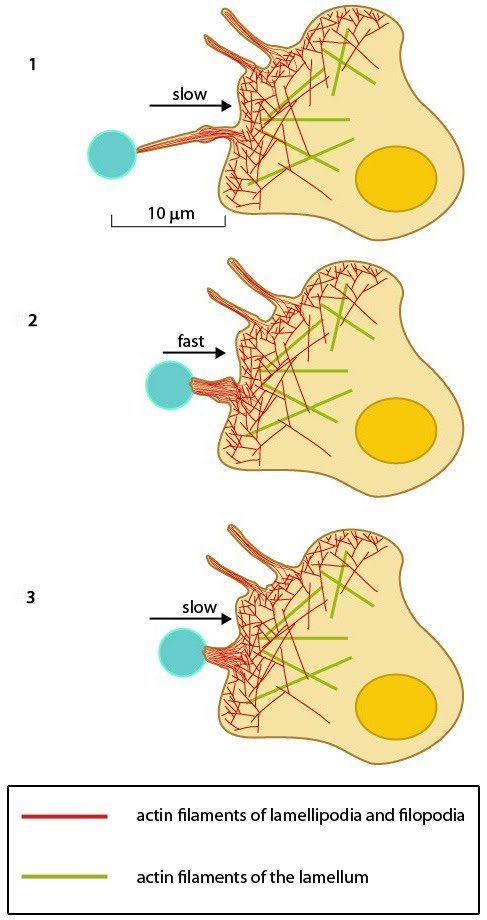 Figure: Filopodia can pull objects. After a filopodium binds to an object, retrograde actin movement and myosin motor activity provide the force(s) needed for pulling the object towards the cell body. Once pulling starts, the initial slow movement (#1) is followed by a burst of rapid movement (#2) that diminishes as the object reaches the cell body (#3) [2]. |
|
|
|
|
MBInfo > Cellular Structures in Mechanosensing and Cell Motility > Filopodia > Steps in Filopodia Formation and Movement >
5. Pulling |
MBInfo © 2013 National University of Singapore.
MBInfo is sponsored by:
Sign in|Report Abuse|Print Page|Remove Access|Powered By Google Sites