
Steps in Formation 1. Initiation 2. Extension 3. Lateral movement and Stasis 4. Adherence 5. Pulling 6. Retraction and Collapse Functional Modules
| Filopodia
|
| System | Maximum Rate | References |
| Mouse cortical neurons; primary mesenchyme cells | 400 nm/s | [4, 5] |
| NG108 | 15 nm/s | [6] |
| Mouse macrophage | 600 nm/s | [7] |
| Chick DRG neurons | 200 nm/s | [8] |
Rapid collapse produces a large number of filopodial strands tightly connected to the substrate by long tethers. F-actin bundles [9] and monomeric actin [2] disappear from collapsing filopodia without a compensatory rise in F-actin at the growth cone center; this indicates a net loss of actin rather than a rearward translocation. Furthermore, active nucleation and protrusion of filopodia is still found in discrete areas of collapsing growth cones, which argues against sequesterization or modification of actin as the mechanism responsible for the loss of F-actin during the collapse [2].
A number of factors regulate collapse and retraction. For example, capping proteins promote filopodial retraction by shielding the barbed end of filaments from further assembly and elongation [10]. Inhibition of F-actin polymerization and protrusion during collapse are mediated by RhoA kinase activity [11]. Collapse may result from the exposure of a “naive” growth cone to a high concentration of a repellent followed by an overactive response [12]. The repulsive component appears to shut down the growth program and is, therefore, dominant over the growth-stimulating effects of adhesion molecules. In addition, the repellent also interferes with mechanisms that would normally result in filopodial retraction [1].
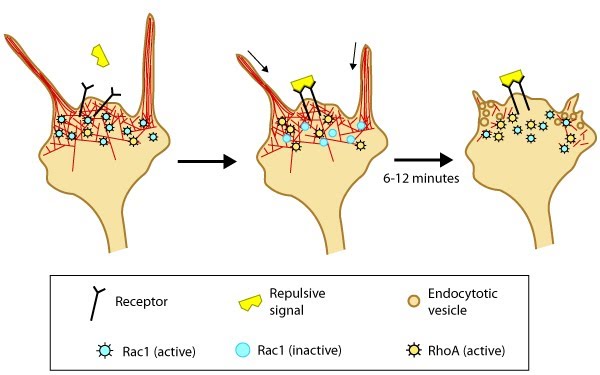 Figure: Model of filopodia collapse. In neuronal growth cones, filopodia protrusion stops when a repulsive signal binds to its receptor on the cell surface. Receptor-binding transiently inactivates Rac1 GTPase and prevents it from promoting actin assembly. Resumption of Rac1 activity coincides with filopodia collapse and is required for endocytosis of the collapsing plasma membrane and reorganization of F-actin [3]. RhoA activity has also been implicated in reducing actin polymerization following treatment with repulsive signals [11].
Figure: Model of filopodia collapse. In neuronal growth cones, filopodia protrusion stops when a repulsive signal binds to its receptor on the cell surface. Receptor-binding transiently inactivates Rac1 GTPase and prevents it from promoting actin assembly. Resumption of Rac1 activity coincides with filopodia collapse and is required for endocytosis of the collapsing plasma membrane and reorganization of F-actin [3]. RhoA activity has also been implicated in reducing actin polymerization following treatment with repulsive signals [11].