Bleb Assembly
Initiation of bleb formation[Edit]
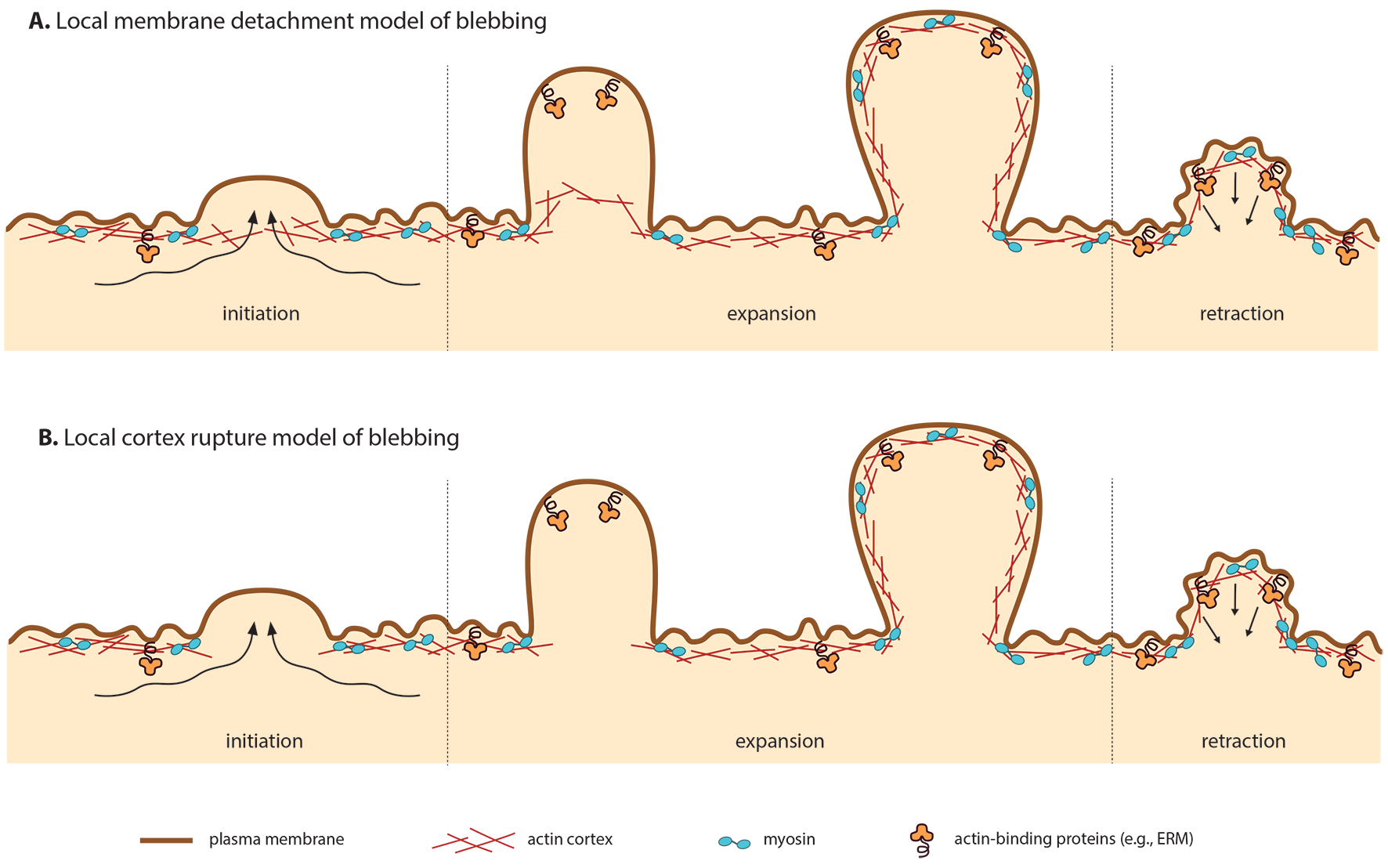
Bleb assembly involves phases of Initiation, Expansion, Repolymerization and Retraction.
Under normal circumstances, the plasma membrane remains tightly bound to the cell cortex. This close association is maintained by interactions with the actin cytoskeleton, myosin and other associated proteins. Hydrostatic pressure is exerted on the plasma membrane via cortical tension generated by myosin [1]. However, during blebbing myosin contracts the cortical actin cytoskeleton, detaching it from the plasma membrane. Depending on whether the cortex remains intact or ruptures during detachment, this process is referred to as local membrane detachment ([2], e.g., zebrafish primordial germ cells [3], Walker carcinoma cells [4]) or local cortex rupture ([5, 6], e.g., Walker carcinosarcona [4]).
Importantly, the initiation of bleb formation is a result of a local increase in hydrostatic pressure, rather than actin
polymerization, as is the case for other cellular structures including lamellipodia and filopodia [7]. The frequency at which blebs are formed is determined by several factors [8], including:
Under normal circumstances, the plasma membrane remains tightly bound to the cell cortex. This close association is maintained by interactions with the actin cytoskeleton, myosin and other associated proteins. Hydrostatic pressure is exerted on the plasma membrane via cortical tension generated by myosin [1]. However, during blebbing myosin contracts the cortical actin cytoskeleton, detaching it from the plasma membrane. Depending on whether the cortex remains intact or ruptures during detachment, this process is referred to as local membrane detachment ([2], e.g., zebrafish primordial germ cells [3], Walker carcinoma cells [4]) or local cortex rupture ([5, 6], e.g., Walker carcinosarcona [4]).
Importantly, the initiation of bleb formation is a result of a local increase in hydrostatic pressure, rather than actin
{kind=link}
polymerization, as is the case for other cellular structures including lamellipodia and filopodia [7]. The frequency at which blebs are formed is determined by several factors [8], including:
- hydrostatic pressure: increased extracellular osmolarity decreases the number of blebs formed
- membrane-cortex adhesion: increased membrane-cortex adhesion decreases the number of blebs formed, as evidenced by the overexpression of the membrane-actin crosslinker ezrin
- membrane tension: increasing membrane tension decreases the number of blebs formed, as evidenced by treating cells with the polysaccharide crosslinker WGA (wheat germ agglutinin)
- cortex contractility: decreased cortical contractility decreases the number of blebs formed, as evidence by treating cells with the myosin II inhibitor, blebbistatin.
Bleb expansion[Edit]
Expansion lasts between 5 to 30 seconds, following bleb initiation and preceding reformation of the cortical actomyosin cytoskeleton just beneath the membrane [9]. During this time, the bleb proceeds to grow as a result of actomyosin driven pressure, resulting in the influx of cytoplasm into the bleb. Cytoplasmic flow within the cell follows a fountain-like pattern through the inner and outer regions [10].
The cytoplasmic volume of the bleb increases in concert with its membrane surface area [11]. Expansion of the membrane surface area is enabled by lipid flow through the neck of the bleb [12,13]. If the growth of the bleb is too rapid to be facilitated by lipid flow alone, the membrane-cortex connection adjacent to the neck of the bleb can get further disrupted due to increased membrane tension [8].
Growing blebs are generally devoid of cortical actin, both in non-motile [2, 6, 14] and motile cells [3, 15, 16]. Instead, they possess a spectrin-based cytoskeleton, such as that observed in red blood cells [17]. The maximal size of the bleb is determined by its initial growth rate, membrane tension and the time required for repolymerization of the cortical cytoskeleton [2].
Motile Blebbing Expansion
Motile blebbing (as opposed to non-motile) occurs primarily at the leading edge. The stimulus and subsequent downstream signaling that initiates this polarization is not yet clear. However, two polarization models have been proposed, one based on the local membrane detachment (1) and the other based on the local cortex ruptre scenario (2) [9]:
Local membrane detachment (1)
In this case, polarization may result from discrete, local weakening of membrane-cortex attachments. This may may be the consequence of uneven distribution of the actin-membrane linker ERM (erzin, radixin and moesin) to the rear of the cell. This model is supported by fixed Walker carcinosarcoma cells [18].
Local cortex rupture (2)
In this case, preferential tearing at the leading edge may result from higher contractility at the rupture point [6], without the need for asymmetry in membrane-cortex attachments.
In both models, old cortex is disassembled at leading edge and new cortex is formed under the growing bleb. Since the leading edge cortex is always younger and more fragile, once the symmetry is broken, polarization continues.
Adherence
In motile blebbing, cells move by exerting a force against the underlying substrate. In lamellipodial motility, this is achieved through adherence of lamellipodia to the substrate. However, in blebbing motility, the mechanism is still unknown. Two models have been proposed [9]:
Weak substrate adhesion (1)
The blebbing cell creates weak adhesions to the substrate. This model is supported by findings from Walker carcinosarcoma cells, showing loose cell-substrate contacts during motile blebbing [19]. Loose cell-cell contacts have also been observed, with contacts spanning a greater distance than that of tight or adherens junctions [20].
Perpendicular force generation (2)
If a motile, blebbing cell is in a confined environment, such as between two glass cover slips, it is able to migrate without the use of cell-substrate adhesions. This is due to the generation of forces perpendicular to the substrate that push the cell forward, squeezing it through the confined space, in a process termed ‘chimneying’ [21]. This is supported by findings in adhesion-deficient leukocytes that are nonetheless able to migrate efficiently [22].
Repolymerization
As expansion of the bleb begins to slow, cortical actin just beneath the bleb membrane begins to repolymerize. This mechanims of actin nucleation is unclear, as two of the most common actin nucleaters, the Arp2/3 complex and the formin mDia1, are not present beneath the membrane of filamin-deficient cells [17]. Studies have, however, shown the sequential recruitment of proteins to the bleb membrane, including F-actin, membrane-linker proteins, actin, actin-bundling proteins, and contractile proteins. The resultant bleb cytoskeleton underlying the membrane comprises multiple layers of F-actin tethered together by actin bundling proteins. During this process of repolymerization in the main cortical body of the bleb, F-actin at the base of the neck of the bleb disassembles due to actin turnover and redistribution [9].
Bleb retraction[Edit]
Retraction generally lasts between 60 to 120 seconds [9]. When the cells membrane lacks the stability afforded, for example, by cell-substrate contacts, the primary cascade involved in retraction is that of mysoin-RhoA-ROCK [23]. In this cascade, myosin is recruited to the bleb lumen, GTP-loaded Rho activates ROCK, which directly phosphorylates myosin light chain and so generates actomyosin contraction [6, 24, 25, 26, 27].
This process can be regulated at multiple levels, both extracellularly and intracellularly, such as in the case of RhoA, its activity can be regulated by both extracellular signals and intracellular cascades [25].
Indirect regulation of this cascade may also occur via Rac GTPases, such as Rac1 [28, 29] or Dictyostelium discoideum RacB [30]. The cascade can also be regulated through inhibition of its effector ROCK [26]. In addition, the formins, Dia1 and mDia2 have been shown to be crucial in regulating retraction [5, 31].
Indirect regulation of this cascade may also occur via Rac GTPases, such as Rac1 [28, 29] or Dictyostelium discoideum RacB [30]. The cascade can also be regulated through inhibition of its effector ROCK [26]. In addition, the formins, Dia1 and mDia2 have been shown to be crucial in regulating retraction [5, 31].
Circus Movement[Edit]
Circus movements have been mainly observed in embryonic blastomeres, neuroectoderm and paraxial mesoderm [32], where a single bleb repeatedly propagates around the circumference of the cell with a period of 1–2 minutes. The cortical actin network reforms asymmetrically during the repolymerization phase of circus movements [8]. This movement requires actomyosin contractility [32, 33, 34].